

Прочетен: 5989 Коментари: 0 Гласове:
Последна промяна: 30.06.2019 22:22
Caenagnathoids (Caenagnathiformes)
 Oviraptorosauria is a clade of Cretaceous theropod dinosaurs of uncertain affinities within Maniraptoriformes.The results of a phylogenetic analysis using 195 characters scored for four outgroup and 13 maniraptoriform (ingroup) terminal taxa, including new data on oviraptorids. This analysis places Oviraptorosauria within Avialae, in a sister-group relationship with Confuciusornis. Archaeopteryx, Therizinosauria, Dromaeosauridae, and Ornithomimosauria are successively more distant outgroups to the Confuciusornis-oviraptorosaur clade. Avimimus and Caudipteryx are successively more closely related to Oviraptoroidea, which contains the sister taxa Caenagnathidae and Oviraptoridae. Within Oviraptoridae, "Oviraptor" mongoliensis and Oviraptor philoceratops are successively more closely related to the Conchoraptor-Ingenia clade. Oviraptorosaurs are hypothesized to be secondarily flightless. Emended phylogenetic definitions are provided for Oviraptoridae, Caenagnathidae, Oviraptoroidea, Oviraptorosauria, Avialae, Eumaniraptora, Maniraptora, and Maniraptoriformes.
Oviraptorosauria is a clade of Cretaceous theropod dinosaurs of uncertain affinities within Maniraptoriformes.The results of a phylogenetic analysis using 195 characters scored for four outgroup and 13 maniraptoriform (ingroup) terminal taxa, including new data on oviraptorids. This analysis places Oviraptorosauria within Avialae, in a sister-group relationship with Confuciusornis. Archaeopteryx, Therizinosauria, Dromaeosauridae, and Ornithomimosauria are successively more distant outgroups to the Confuciusornis-oviraptorosaur clade. Avimimus and Caudipteryx are successively more closely related to Oviraptoroidea, which contains the sister taxa Caenagnathidae and Oviraptoridae. Within Oviraptoridae, "Oviraptor" mongoliensis and Oviraptor philoceratops are successively more closely related to the Conchoraptor-Ingenia clade. Oviraptorosaurs are hypothesized to be secondarily flightless. Emended phylogenetic definitions are provided for Oviraptoridae, Caenagnathidae, Oviraptoroidea, Oviraptorosauria, Avialae, Eumaniraptora, Maniraptora, and Maniraptoriformes.

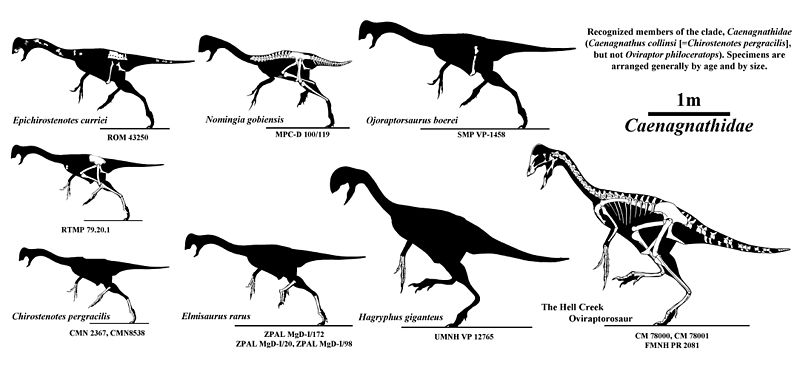
1. Gigantoraptor 2. Nomingia 3. Oviraptor 4. Chirostenotes 5. Caudipteryx
Caenagnathoids ranged in size from Caudipteryx, which was the size of a turkey, to the 8 metre long, 1.4 ton Gigantoraptor.
The earliest and most basal ("primitive") known oviraptorosaurs are Ningyuansaurus wangi, Protarchaeopteryx robusta and Incisivosaurus gauthieri, both from the lower Yixian Formation of China, dating to about 125 million years ago during the Aptian age of the early Cretaceous period. A tiny neck vertebra reported from the Wadhurst Clay Formation of England shares some features in common with oviraptorosaurs, and may represent an earlier occurrence of this group (at about 140 million years ago).
Caenagnathoids like deinonychosaurs, are so bird-like that several scientists consider them to be true birds, more advanced than Archaeopteryx. Gregory S. Paul has written extensively on this possibility, and Teresa Maryańska and colleagues published a technical paper detailing this idea in 2002.Michael Benton, in his widely respected text Vertebrate Paleontology, also included oviraptorosaurs as an order within the class Aves.However, a number of researchers have disagreed with this classification, retaining oviraptorosaurs as non-avialan maniraptorans slightly more primitive than the deinonychosaurs.
Largest Caenagnathoids was Gigantoraptor erlianensis
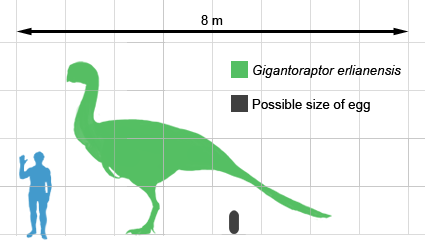
Gigantoraptor is a genus of giant oviraptorosaurian theropod.It was discovered in 2005 in the Iren Dabasu Formation, Erlian basin, in Inner Mongolia. The age of the Iren Dabasu formation is controversial. Based on ostracods, Godefroit suggested the unit was Nemegt equivalent or about 70 million years old, although some dinosaur remains suggest an older Santonian age c 84-86 Ma.
In a quarry at Saihangaobi, in Sonid Left Banner (Inner Mongolia), numerous remains of the sauropod Sonidosaurus have been uncovered since 2001. Chinese paleontologist Xu Xing was asked to reenact the discovery of Sonidosaurus in April 2005 for a Japanese documentary. Xu obliged them by digging out a thighbone. As he wiped the bone clean, he suddenly realized it was not from a sauropod, but from an unidentified theropod in the size class of Albertosaurus. He then stopped the filming to secure the serendipitous find. This way, the discovery of the Gigantoraptor holotype fossil was documented on film.
In 2007, the type species Gigantoraptor erlianensis was named and described by Xu, Tan Qingwei, Wang Jianmin, Zhao Xijin and Tan Lin. The generic name is derived from Latin gigas, gigantis, "giant" and raptor, "seizer". The specific name refers to the Erlian Basin.
The only known type specimen, LH V0011, was found in layers dating to the Santonian, about eighty five million years old. It consists of is the incomplete and disassociated remains of a single subadult individual, a partial skeleton lacking the cranium but including the lower jaw, a single neck vertebra, most of the back and tail and the majority of the frontlimb and hindlimb elements.
It was clear to Xu et al. that Gigantoraptor belonged to the Oviraptorosauria, a group named after Oviraptor, but compared to other known members, Gigantoraptor was much larger, approximately three times as long and 35 times more massive than the heaviest earlier discovered oviraptorosaurid Citipati. Xu et al. estimated the length at 8 metres (26 ft) and the weight at 1400 kilogrammes. In 2010, Gregory S. Paul even gave an estimate of two tonnes (2.2 tons).
The toothless lower jaws of Gigantoraptor are fused into a broad shovel-like mandibula. They indicate that the unknown skull was over half a metre long and toothless also, probably equipped with a horny beak. The front tail vertebrae have very long neural spines and are heavily pneumaticised with deep pleurocoels. The middle section of the relatively short tail is somewhat stiffened by long prezygapophyses. The back tail vertebrae are lightened by spongeous bone. The front limb is rather long because of an elongated slender hand. The humerus is bowed outwards to an exceptionally large extent and has a very rounded head. The first metacarpal is very short and carries a strongly diverging thumb. The hindlimb is also long because of an elongated lower leg. The thighbone is relatively slender and short with a distinct head and neck. The foot is robust with large and strongly curved toe claws.
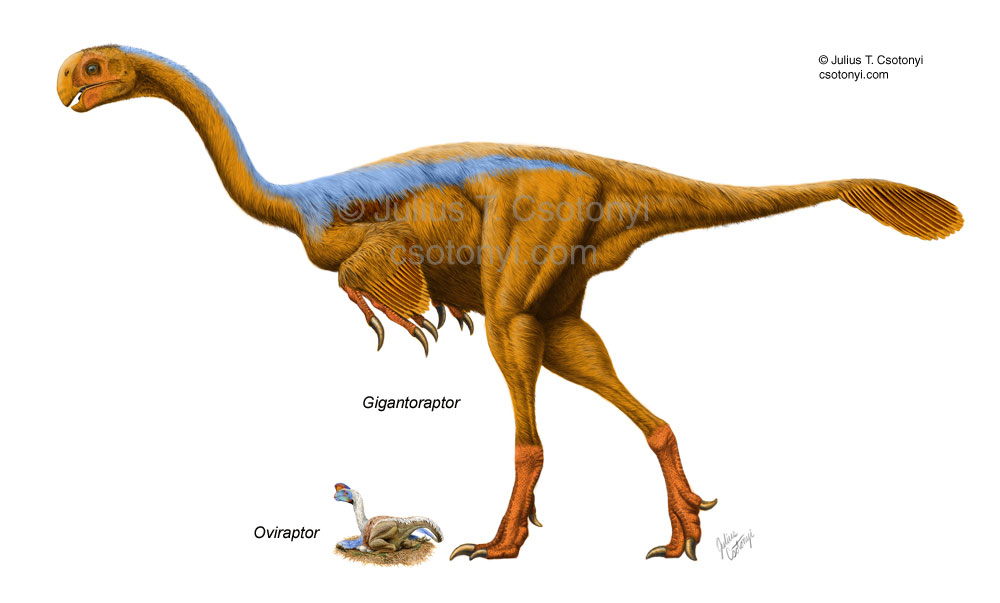
No direct evidence of feathers was preserved with the skeleton, but Xu et al. (2007) discussed their likely presence on Gigantoraptor. They admitted that despite Gigantoraptor being a member of the Oviraptorosauria, a group that includes the feathered species Caudipteryx and Protarchaeopteryx, it might have been "naked" because it is three hundred times as massive as these species, and very large animals may rely more on mass for temperature regulation, losing the insulating coverings found on their smaller relatives. However, they suggested that at least arm feathers were probably still present on Gigantoraptor, since their primary functions, such as display and covering the eggs while brooding, are not related to the regulation of body heat.
Eumaniraptorans & Deinonychosaurians(Dromaeosauridae)
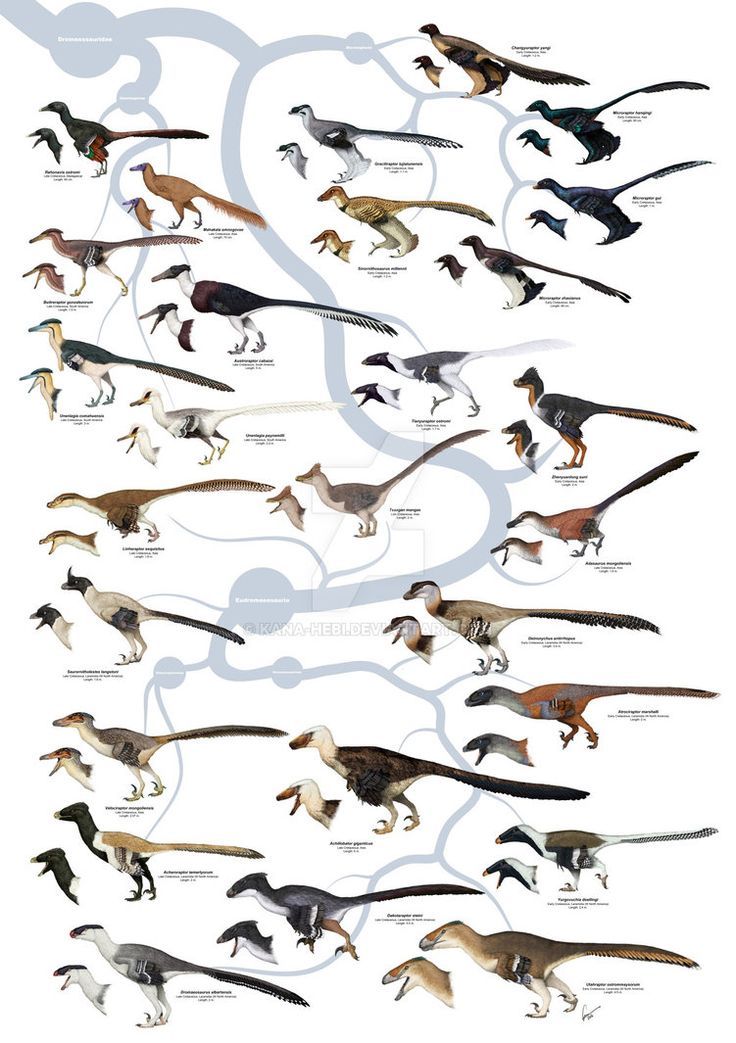
Dromaeosaurs constitute a small clade of theropod dinosaurs which exhibit some highly derived characteristics that they all share, especially modifications of the forelimb allowing for a flexible seizing function (which is thought to have been modified to create the bird "flight stroke"). According to current thinking, birds are hypothesized to have shared a common ancestor with the dromaeosaurs sometime in the Jurassic period; Dromaeosauridae is thus termed the sister group of the clade Aves (which includes all birds). Dromaeosaurid fossils have been found across the globe in North America, Europe, Africa, Asia, South America and Antarctica,with fossilized teeth giving credence to the possibility that they inhabited Australia as well.They first appeared in the mid-Jurassic Period (late Bathonian stage, about 167 million years ago) and survived until the end of the Cretaceous (Maastrichtian stage, 66 ma), existing until the Cretaceous–Paleogene extinction event. The presence of dromaeosaurids as early as the Middle Jurassic has been suggested by the discovery of isolated fossil teeth, though no dromaeosaurid body fossils have been found from this period.
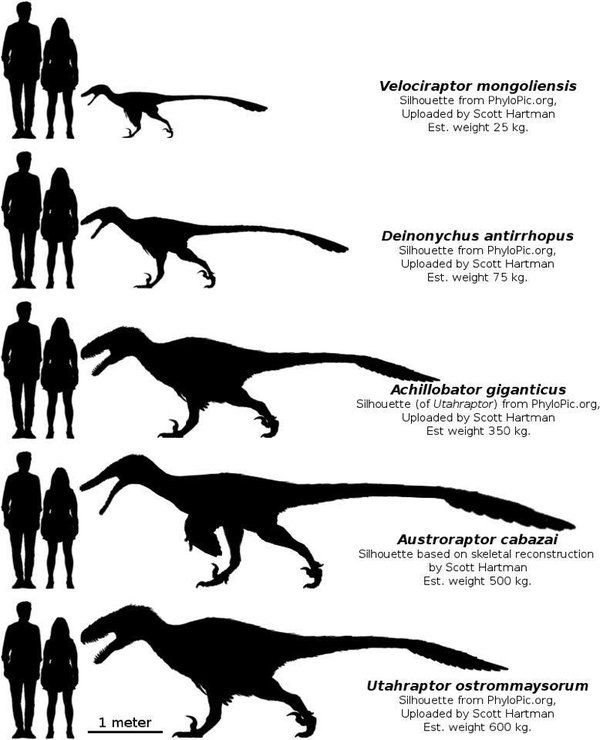
Dromaeosaurids were small to medium-sized dinosaurs, ranging from about 0.7 metres (2.3 ft) in length (in the case of Mahakala) to approaching or over 6 m (20 ft) (in Utahraptor, Dakotaraptor and Achillobator).Some may have grown larger; undescribed specimens of Utahraptor in Brigham Young University collections belonged to individuals that may have reached up to 11 m (36 ft) long, though these await more detailed study.Large size appears to have evolved at least twice among dromaeosaurids; once among the dromaeosaurines Utahraptor and Achillobator, and again among the unenlagiines (Austroraptor, which measured 5 m [16 ft] long).
A possible third lineage of giant dromaeosaurids is represented by isolated teeth found on the Isle of Wight, England. The teeth belong to an animal the size of the dromaeosaurine Utahraptor, but they appear to belong to velociraptorines, judging by the shape of the teeth. Mahakala is both the most primitive dromaeosaurid ever described and the smallest. This evidence, combined with the small size of other primitive relatives such as Microraptor and the troodontid Anchiornis, indicates that the common ancestor of dromaeosaurids, troodontids, and birds (which is called the ancestral paravian) may have been very small, at around 65 cm in length and 600 to 700 grams of mass.

Dromaeosaurids share many features with early birds (clade Avialae or Aves). The precise nature of their relationship to birds has undergone a great deal of study, and hypotheses about that relationship have changed as large amounts of new evidence became available. As late as 2001, Mark Norell and colleagues analyzed a large survey of coelurosaur fossils and produced the tentative result that dromaeosaurids were most closely related to birds, with troodontids as a more distant outgroup. They even suggested that Dromaeosauridae could be paraphyletic relative to Avialae.In 2002, Hwang and colleagues utilized the work of Norell et al., including new characters and better fossil evidence, to determine that birds (avialans) were better thought of as cousins to the dromaeosaurids and troodontids.A consensus of paleontologists has concluded that there is not yet enough evidence to determine whether any dromaeosaurids could fly or glide, or whether they evolved from ancestors that could.
Largest dromaeosaurid was Eudromaeosaurid Utahraptor
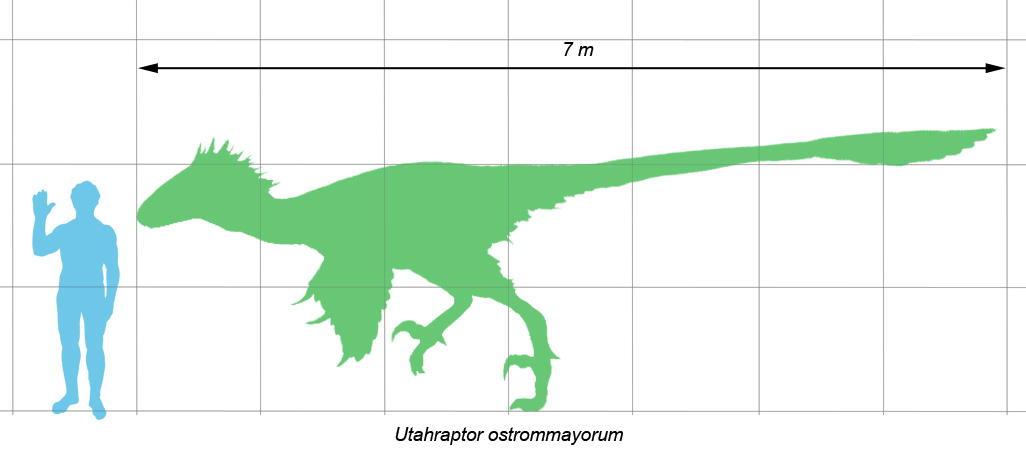
Utahraptor (meaning "Utah"s predator) is a genus of dromaeosaurid theropods . It contains a single species, Utahraptor ostrommaysorum, which is the largest known member of the family Dromaeosauridae. Fossil specimens date to the upper Barremian stage of the early Cretaceous period (in rock strata dated to 126 ± 2.5 million years ago).
The holotype specimen of Utahraptor is fragmentary, consisting of skull fragments, a tibia, claws and some caudal (tail) vertebrae. These few elements suggest an animal about twice the length of Deinonychus.Like other dromaeosaurids, Utahraptor had large curved claws on their second toes. One claw specimen is preserved at 22 centimetres (8.7 in) in length and is thought to reach 24 centimetres (9.4 in) restored.
The largest described U. ostrommaysorum specimens are estimated to have reached up to 7 m (23 ft) long and somewhat less than 500 kg (1,100 lb) in weight, comparable to a polar bear in size.
It is thought that Utahraptor may be closely related to the smaller Dromaeosaurus and the giant Mongolian dromaeosaurid genus Achillobator.
Although feathers have never been found in association with Utahraptor specimens, there is strong phylogenetic evidence suggesting that all dromaeosaurids possessed them. This evidence comes from phylogenetic bracketing, which allows paleontologists to infer traits that exist in a clade based on the existence of that trait in a more basal form. The genus Microraptor is one of the oldest known dromaeosaurids, and is phylogenetically more primitive than Utahraptor. Since Microraptor and other dromaeosaurids possessed feathers, it is reasonable to assume that this trait was present in all of Dromaeosauridae. Feathers were very unlikely to have evolved more than once, so assuming that any given dromaeosaurid, such as Utahraptor, lacked feathers would require positive evidence that they did not have them. So far, there is nothing to suggest that feathers were lost in larger, more derived species of dromaeosaurs.
In a 2001 study conducted by Bruce Rothschild and other paleontologists, two foot bones referred to Utahraptor were examined for signs of stress fracture, but none were found.
The first specimens of Utahraptor were found in 1975 by Jim Jensen in the Dalton Wells Quarry in east-central Utah, near the town of Moab, but did not receive much attention. After a find of a large foot-claw by Carl Limoni in October 1991 James Kirkland, Robert Gaston, and Donald Burge uncovered further remains of Utahraptor in 1991 in the Gaston Quarry in Grand County, Utah, within the Yellow Cat and Poison Strip members of the Cedar Mountain Formation.Radiometric dating has shown that these parts of the Cedar Mountain Formation were deposited about 124 million years ago.The type specimen, CEU 184v.86, is currently housed at the College of Eastern Utah Prehistoric Museum, although Brigham Young University, the depository of Jensen"s finds, currently houses the largest collection of Utahraptor fossils.
The type species (and only known species of Utahraptor), Utahraptor ostrommaysorum, was named by Kirkland, Gaston, and Burge in June of 1993 for the American paleontologist John Ostrom from Yale University"s Peabody Museum of Natural History, and Chris Mays of Dinamation International. Originally, in the specific name, the singular genitive ostrommaysi was used but, in 2000, this was emended by George Olshevsky to the plural.Earlier, it had been intended to name the species "Utahraptor spielbergi", after film director Steven Spielberg, in exchange for funding palaeontological research, but no agreement could be reached on the amount of financial assistance.
In 2001, Kirkland et al. discovered a fossil block of sandstone in eastern Utah containing as many as six individuals, including an adult measuring about 4.8 m (16 ft), four juveniles and a baby about 1 m (3.3 ft). Also fossilized with the predators are the remains of an iguanodont. It is theorized by Kirkland that the Utahraptors attempted to attack helpless prey mired in thick mud or quicksand, and were themselves mired in the attempt to attack the victim. Similar sites such as the Cleveland-Lloyd Quarry and La Brea Tar pits house similar predator traps. Examination of the fossil is ongoing after a decade of excavation, but if correct it may be one of the best preserved predator traps ever discovered. The fossil may further reveal aspects into the behavior of Utahraptor, such as whether it might have hunted in groups like Deinonychus is believed to have done.
Achillobator giganticus
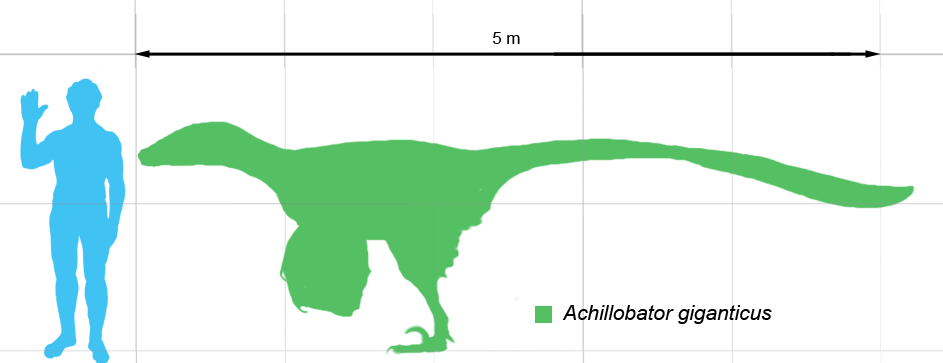
Achillobator is a dromaeosaurid theropod that lived roughly 98 to 83 million years ago during the Late Cretaceous in what is now Mongolia, in Asia. It was among the largest dromaeosaurs; the holotype and only known individual of Achillobator is estimated at 5 to 6 m (16.4 to 19.7 ft) long. Achillobator was a moderately-built, ground-dwelling, bipedal carnivore. It would have been an active predator, hunting with the enlarged, sickle-shaped claw on the second toe.
Fossils of Achillobator were first discovered during a Mongolian and Russian field expedition, and collected by Burkhant in 1989, but the specimen was not described and named until 1999 by Mongolian paleontologist Altangerel Perle, and American paleontologists Mark Norell and Jim Clark, although the description was not complete and was actually published without the knowledge of the latter two paleontologists.
The fossils of the type specimen of Achillobator, FR.MNUFR 15, were found associated but mostly disarticulated, and include a left maxilla with teeth, two cervical vertebrae, two dorsal vertebrae, rib fragments, seven caudal vertebrae, a scapula and coracoid, a pelvis with a right ilium, pubis and ischium, a radius, an incomplete manus, a left femur and tibia, and an incomplete pes. Smith at al. (2012) noted that this genus represents the second largest of the known dromaeosaurid taxa with a tibial length of 490 mm. Its femur, which is 3% longer than the tibia, a rare trait in Dromaeosaurs, measures 505 mm in length. Estimates suggest that Achillobator weighed 350 kg (771.6 lb) at most. The teeth are serrated and recurved, and the posterior serrations are slightly larger than the anterior serrations.
The genus name Achillobator means "Achilles hero" and is derived from Achilles, a famous ancient Greek warrior who fought in the Trojan War, and the Mongolian word baatar, anciently bagatur, which means "hero". The generic name refers to the large Achilles tendon that connects to the sickle claw on the foot, which was the major combat weapon of dromaeosaurids.
The pelvis of Achillobator seems to show plesiomorphic ("primitive") saurischian characteristics compared to other dromaeosaurids. For instance, the pubis is aligned vertically and has a large pubic boot (a wide expansion at the end), unlike most other dromaeosaurids, where there is a much smaller boot, if any, and the pubis points backwards in the same direction as the ischium (a condition called opisthopuby, which is also seen in the unrelated therizinosaurs and ornithischians, as well as in birds).
The above differences and others led Burnham et al. (2000) to suggest that Achillobator represents a paleontological chimera. However, other studies have attempted to refute this, noting that many pieces were found to be semi-articulated, all of the elements are the same color and preservation, and that Achillobator routinely comes out as a dromaeosaurid in cladistic analyses, even taking into account the differences.
The remains of Achillobator were recovered in the Bayan Shireh Formation of Dornogovi Province, Mongolia, in fine-grained, medium sandstone/gray mudstone that was deposited during the Late Cretaceous epoch. The exact age is uncertain, with two competing hypotheses; based on comparisons with other formations, the Bayan Shireh fauna seems to correspond best with the Turonian through early Campanian stages of the Late Cretaceous, about 93 to 80 million years ago. However, examination of the magnetostratigraphy of the formation seems to confirm that the entire Bayan Shireh lies within the Cretaceous Long Normal, which lasted only until the end of the Santonian stage, giving a possible Cenomanian through Santonian age, or between 98 and 83 million years ago.
The paleofauna of the Bayan Shireh Formation included, the therizinosaurid Segnosaurus, the deinocheirid Garudimimus, and the ankylosaurid Talarurus, as well as a genus of sauropod, a tyrannosauroid formerly referred to Alectrosaurus, and an azhdarchid pterosaur.
Dakotaraptor
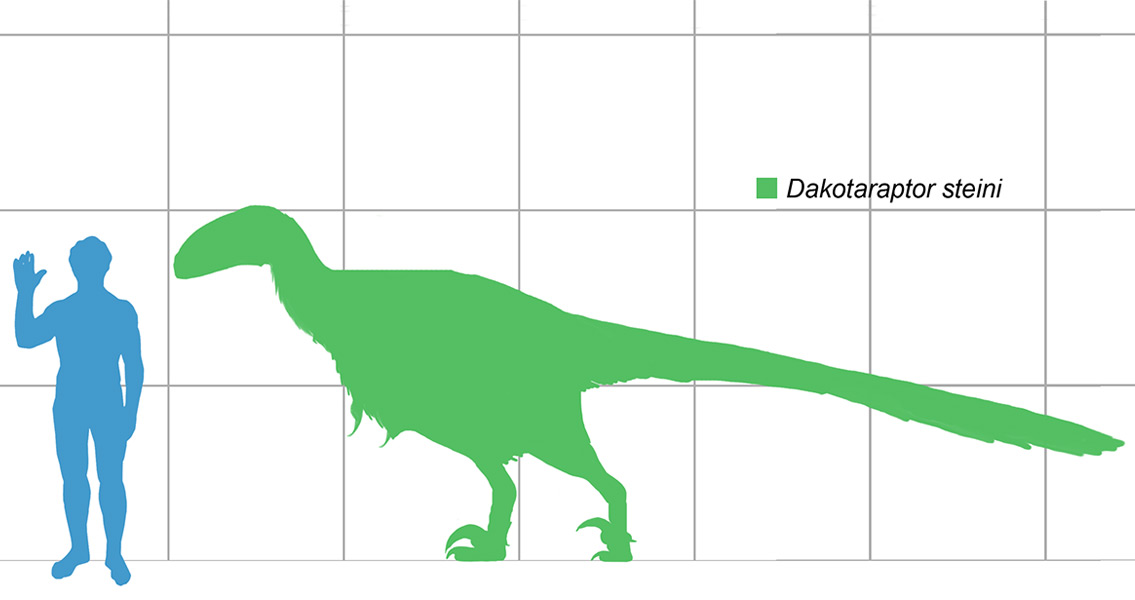
The one thing that immediately stands out about Dakotaraptor at first glance is simply the large size of this dromaeosaurid. Most dromaeosaurids measure in at under two meters in length, with a few genera ranging between two and three meters long. The Holotype individual of Dakotaraptor however has a reconstructed length of five and a half meters. At the time of the 2015 description, this makes Dakotaraptor the second largest dromaeosaurids known, with the largest being Utahraptor.
As far as comparison between Dakotaraptor and Utahraptor goes, Utahraptor lived in North America around Utah (clue is in the name) during the Early Cretaceous roughly one hundred and twenty-six million years ago. Dakotaraptor also lived in North America but around South Dakota towards the end of the Late Cretaceous some sixty-six million years ago. Not only are Dakotaraptor and Utahraptor separated by a wide temporal gulf, study of the known fossils of Dakotaraptor show that in body proportions this genus was actually more like genera such as Deinonychus and Dromaeosaurus than it was Utahraptor, making a direct link between Dakotaraptor and Utahraptor unlikely.
Studies of dromaeosaurid dinosaurs going back to the late twentieth century indicate that dromaeosaurid dinosaurs almost certainly had feathers. These feathers were not necessarily all over the body, but were likely on the thorax and abdomen and almost certain on the arms. Dakotaraptor supports this theory in that fossils of the ulna (one of the forearm bones) show attachment points for large pennaceous feathers. It is unknown to what extent feathers would have covered Dakotaraptor, but smaller insulating feathers are known to have covered the body in even larger theropod dinosaurs such as the tyrannosaur Yutyrannus. However other dinosaurs living in North America during the late Cretaceous such as the ornithomimosaur Ornithomimus are known to have had small feathers on the back and sides of the body, but have legs that were devoid of feathers. The coverage of feathers on Dakotaraptor were likely a reflection of environmental factors first, and additional functions such as display and egg insulation second.
In the early twenty-first century there has been a lot of debate concerning the relationship between dromaeosaurid dinosaurs and the evolution of birds, with the classic idea being that birds evolved from maniraptoran dinosaurs. There are now alternate theories that maniraptoran dinosaurs may have evolved from birds, or perhaps even to birds and back again, but the ideas are simply too long to explain and off topic for this article. The key point is that some dromaeosaurs may have had a limited flight ability even if only gliding as has been speculated for genera such as Microraptor. As far as Dakotaraptor is concerned, flight was almost certainly an impossibility, mainly for the simple fact that at about five and a half meters long Dakotaraptor would have simply been too big. There was also no way for Dakotaraptor to use its arms to generate anywhere near enough lift. The original study of the forearms of Dakotaraptor into wings, indicated the total ‘wingspan’ of Dakotaraptor would have been about one hundred and twenty centimetres, which is even less than what an average human adult could achieve if they held their arms out to the side.
Although certainly too big to fly, Dakotaraptor was still no slouch on the ground. Proportions of the leg show that the lower leg bones of the tibia and fibula were longer than the femur which makes up the upper leg. This allows for a significantly longer stride of the leg which directly translates to an increase in speed and ability to run fast. The vertebrae of Dakotaraptor were also pneumatised, meaning that they had air spaces within them. This dramatically reduces the weight of the bone, and with this factor in mind, Dakotaraptor would have actually been quite lightweight for its size. Not only would this allow for larger size growth, but it would also allow Dakotaraptor to retain some of the speed and agility that dromaeosaurid dinosaurs are known for.
At the original fossil site of Dakotaraptor, remains of more than one individual were found. The Holotype individual is the most complete of these, however there also seems to have been a notable difference in build between the holotype which is by far the most robust (heavily built), and the others which are more gracile (lightly built). All individuals seem to have been fully grown adults, so one train of thought is that these may represent a collection of male and female individuals. However that still does not make it certain which is which, as while in the animal kingdom males are usually the more robustly built, it is not unknown for females to have been the larger (the moa bird Dinornis is a good example). Only further study and fossil remains can tell us for sure.
Austroraptor cabazai

Austroraptor is an extinct genus of dromaeosaurids that lived about 70 million years ago during the Cretaceous Period in what is now modern Argentina. Austroraptor was a medium sized, moderately-built, ground-dwelling, bipedal carnivore, that could grow up to 5 m (16.4 ft) long. Its length makes Austroraptor one of the largest dromaeosaurids known, with only Achillobator, Dakotaraptor, and Utahraptor approaching or surpassing it in length. It is the largest dromaeosaur to be discovered in the Southern Hemisphere. Particularly notable about the taxon were its relatively short forearms, much shorter in proportion when compared to the majority of the members of its family.
Considered large for a dromaeosaur, Austroraptor cabazai measured around 5 metres (16.4 ft) in length from head to tail. It is the largest dromaeosaur to be discovered in the Southern Hemisphere.The type specimen, labeled MML-195, consisted of a fragmentary skeleton including parts of the dinosaur"s skull, a few neck and torso vertebrae, some ribs, a humerus, and assorted bones from both legs. However little of the entire skeleton was found, what bones were available for analysis expressed some distinct characteristics that differentiate Austroraptor from other dromaeosaurs. A. cabazai"s 80 centimeter-long skull was low and elongated, much more so than that of other dromaeosaurs. Several of its skull bones bore some resemblance to that of the smaller troodontid deinonychosaurs. The front limbs of this taxon were short for a dromaeosaur, with its humerus less than half the length of its femur.Among the Dromaeodauridae, only this genus, Tianyuraptor and Mahakala have similarly reduced forelimbs.The relative length of its arms has caused Austroraptor to be compared to another, more famous short-armed dinosaur, Tyrannosaurus.
Austroraptor had conical, non-serrated teeth, which Novas et al. compared to those of spinosaurids, based on how the enamel of the surface of its teeth is fluted.Austroraptor shares a trait that is unique to it and to Adasaurus mongoliensis: the descending process of the lacrimal bones curves anteriorly to a large degree.Austroraptor has a bizarre morphology in its pedal phalanges, which are strangely disproportionate. Phalanx IV-2 is over twice the width of phalanx II-2, and nearly three times the expected width based on similarly sized members of its taxonomic family.This has suggested to some researchers that the holotype specimen is a chimera, which means that its bones may be from more than one individual.
A diagnosis is a statement of the anatomical features of an organism (or group) that collectively distinguish it from all other organisms. Some, but not all, of the features in a diagnosis are also autapomorphies. An autapomorphy is a distinctive anatomical feature that is unique to a given organism or group. According to Novas et al. (2009), Austroraptor can be distinguished based on the following characteristics: a lacrimal that is highly pneumatized, with the descending process strongly curved rostrally*, and with a caudal process flaring out horizontally above the orbit, the postorbital bone is lacking a dorsomedial process for articulation with the frontal bone*, and with the squamosal process extremely reduced, the maxillary and dentary teeth are small, conical, devoid of serrations and fluted, the humerus is short, representing slightly less than 50 per cent of length of the femur, the pedal phalanx II-2 is transversely narrow, contrasting with the extremely robust phalanx IV-2.
The type specimen of Austroraptor cabazai was recovered in the Bajo de Santa Rosa locality of the Allen Formation, in Rнo Negro, Argentina. The specimen was collected by Phil Currie & Paulina Carabajal in 2008. This specimen is housed in the collection of Museo Municipal de Lamarque in Argentina.
The genus name Austroraptor, means "Southern thief", and is derived from the Latin word auster meaning "the south wind", and the Latin word raptor meaning "thief". The specific name cabazai, was named in honor of Alberto Cabaza, who founded the Museo Municipal de Lamarque where the specimen was partially studied.Austroraptor was described and named by late 2008 by Fernando Novas of the Museo Argentino de Ciencias Naturales. The type species is Austroraptor cabazai.
A cladistic analysis of the specimen"s anatomical features by the describers placed Austroraptor within the subfamily Unenlagiinae of the Dromaeosauridae.This assignment was based on characteristics observed in the bones of the skull, the teeth, and the geometry and formation of the specimen"s vertebral elements. It was determined that Austroraptor was a close relative of the unenlagiine dromaeosaur Buitreraptor, with which it shares certain derived characteristics of the neck vertebrae.
The specimen was found in terrestrial sediments that were deposited during the Maastrichtian stage of the Cretaceous period, approximately 70 million years ago.
Austroraptor shared its paleoenvironment with early mammals, pterosaurs and the titanosaurids Saltasaurus and Rocasaurus, which may have served as prey for this dromaeosaur.
Тагове:
Encyclopedia Largest prehistoric animals...
Encyclopedia Largest prehistoric animals...
