

Прочетен: 9678 Коментари: 0 Гласове:
Последна промяна: 12.08.2022 19:13
The rhinocerotoids of the early Eocene and subsequently the Hyracodontidae and Amynodontidae show no sign of dermal armour or horns. The oldest known genus of rhinocerotids, or true rhinoceroses, is Teletaceras from the Middle Eocene of North America and the Late Eocene of Asia. Due to the timing of the land bridges, it does not appear in South America.It features the first known rhinocerotid tusks, a derived feature, which are still extant in three species and are "the primary offensive weapons" of those species.They appear to have been sexually dimorphic and has features convergent with the development of tusks in pigs, hippopotamuses and elephant seals.
The first dermal armour is indicated by cranial rugosity on Trigonias osborni, Late Eocene, 42-32 Mya. It appears more developed in Subhyracodon, early Oligocene, 33 Mya, both of North America.There appears to have been a correlation between the tusks and the armour later (4 Mya later) defending against them.

Having begun to diversify (increase in species and genera) in America, the rhinocerotids spread rapidly over Beringia when that bridge appeared in the Early Oligocene to Eurasia, where subsequently they became highly diverse. The family reached a maximum floruit in the Miocene. At that time, they entered Africa, but never were very diverse there. In the Pliocene they declined, disappearing from America. The Pleistocene brought extinction from Eurasia, but African and Southeast Asian species survived.
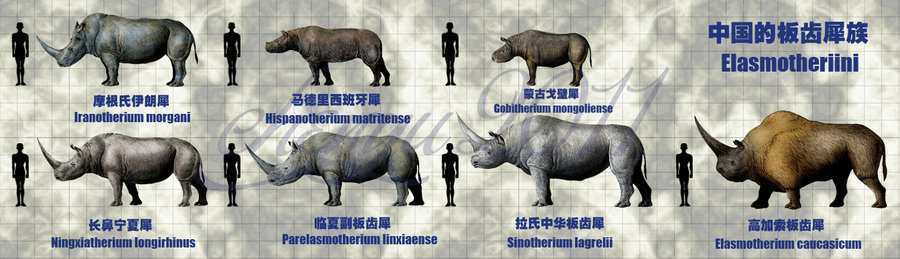
The classification of all the genera and species of the rhinoceros family below the level of Rhinoceratidae has changed more rapidly in the first decade of the 21st century due to, first, the continued discovery of fossils representing new species and genera from China and Mongolia, and second, the number of points of morphology to be compared in computerised phylogenetic analysis. Two branches are generally recognised that lead to the smaller rhinoceroses and the Pliocene/Pleistocene branch of the larger elasmotheres. Both began from small species in the steppes of the Far East, but those in turn are comparable to North American species. Whether the branch point is considered to be at the subfamily level (Elasmotheriinae versus Rhinocerotinae) or at the tribe level (Elasmotheriini versus Rhinocerotini, the currently accepted version) or whether the elasmotheres existed at the subtribe level (Elasmotheriina, a discontinued taxon) is a matter of the status quo in publications, fossils and software programs.
In 2002 P. O. Antoine performed a cladistic analysis using 282 "cranial, dental and postcranial characters"of 28 "terminal taxa" of "elasmotheriines from China and Mongolia" and four outgroups. He found that the elasmotheres were a monophyletic group.
The main characters of derived elasmotheres (huge size, frontal horn, ossified nasal septum, enamel folding, hypsodonty, loss of anterior dentition, lengthening of the molar series ...) are absent in Middle Miocene Elasmotheriina from China and Mongolia.Most of these features appear somewhat later, during the Late Miocene or the Pliocene, in Parelasmotherium, Sinotherium and then Elasmotherium.
Antoine was following the now-discontinued practice of considering the Miocene ancestral species as elasmotheres. They were not, however, distinguished by the features of elasmotheres. The original ancestors were "minute brachydont animals". Elasmotherium was a "mammoth-sized hypsodont".
Such cranio-dental evolution demonstrates unequivocally the increasing proportion of grass-eating in the elasmotherine diet throughout the Middle Miocene.
Parelasmotherium

Parelasmotherium is an extinct genus of rhinos that lived in northern late China about 11 million years ago in northern China. It belonged to the group of Elasmotheriini with his large physique and his hochkronigen teeth and was thus a relative of the Elasmotherium spread in the Pleistocene over much of northern Asia.
Parelasmotherium was a large representative of rhinos, but so far is largely known only from isolated teeth, individual skeletal elements of the musculoskeletal system and a complete, but laterally slightly depressed skull. This was extremely elongated at 97 cm and very narrow. The occipital bone was slightly withdrawn and designed rather rectangular and had a clear, straight in the plan just extending bead as a point of attachment for the neck muscles. The long outstretched nose bone showed a strong build and showed a clear curve in the front part. In contrast to later Elasmotherien had the nasal septum still no ossified structures. Pearl-shaped roughening on the surface of the nasal bone marked the point of attachment for the horn, which was not sitting directly on the tip of the nose, but rather in the middle area of the nasal bone. A forehead horn as in Elasmotherium was not formed. The eye socket was located above behind the last molar.
The maxillary dentition was characterized by a significant reduction of the anterior dentition as with all Elasmotheriini, so that the incisors and the canine were missing. The posterior dentition consisted of three premolars and three molars, in the deciduous dentition was also the foremost premolar. The premolars were smaller than the molars, but resembled these in construction. In general, the teeth showed very high dental crowns, the second molar, which was also the largest tooth in the dentition at the same time, was up to 13 cm high. Here, the values for the height of the molars were higher than those of the length and thus refer to the later representatives of Elasmotherien with their even higher tooth crowns, while in the earlier genera, the height values did not reach the length. In addition, they had a high proportion of cement and significantly folded enamel, but had not so distinctive curves, as in the later Elasmotherium. The high tooth crowns and the construction of the teeth suggest that Parelasmotherium was a specialized grass eater (grazer), which corresponds to the late Miocene climate and associated landscape changes in the form of the spread of open steppes.
Parelasmotherium belongs to the tribe of Elasmotheriini, a group of very large rhinos with very high-crowned molars, which represent the closest relatives of Rhinocerotini with the rhino species still living today. Within the Elasmotheriini the genus forms a clade with Ningxiatherium. Both rhinoceros forms are assigned together with the outermost Iranotherium due to the location of their horn on the nose and the generally not so high-crowned molars and less pleated enamel of the sub-tribe Iranotheriina, the tribally younger frontal horn-bearing and extremely hochkronigen modern Elasmotheiina, which includes the known Elasmotherium heard face. Some researchers also put Ningxiatherium in the genus Parelasmotherium, but with the discovery of the first complete Parelasmotherium skull early in the 21st century, there were marked differences between the two genera, including the lack of ossification of the anterior part of the Nasal septum and the differently built occipital bone exist at Parelasmotherium.
Ningxiatherium

Ningxiatherium is an extinct genus of very large rhinos that lived in northern China about 11 million years ago in the Miocene. It belongs to the group of Elasmotheriini, which is generally characterized by its large body size and high tooth crowns of the molars with partly clearly folded tooth enamel and represents the nearest related group to the today living rhino species.
Ningxiatherium representatives were very large rhinos. The genus is known from two largely complete skull findings and few isolated teeth. The skull was elongated and narrow and reached a length of 93 to 100 cm. The elongated and acute-angled occipital bone caused a relatively low head posture. The nasal bone was rounded and had an obstructed nasal septum in the anterior third. The frontal line between the nasal and occipital bones showed only slight saddling. Rough, speckled surface structures on the top of the skull indicated the position of the horn, which did not sit on the forehead but on the nasal bone, as in the later Elasmotherias. A frontal horn was not formed at Ningxiatherium. The location of the orbit was at the level of the end of the last molar and thus very far back in the skull.
The maxillary dentition was significantly reduced and consisted of at least three premolars and three molars, incisors and canines were not formed. But that was a bit more extensive than that of his later descendant, such as the Elasmotherium. The two rows of teeth were clearly parallel to each other. The anterior premolars were relatively small, while the latter almost reached the dimensions of the next molar and was also characteristically molarized. Largest, however, was the second molar. As with all Elasmotheriini, the teeth were extremely high-crowned (hypsodont), but the enamel did not show such strong folds.
Ningxiatherium belongs to the tribe of the Elasmotheriini, which forms a subgroup of the Rhinocerotidae and the sister taxon of the Rhinocerotini, which include today"s rhinos. Within the Elasmotheriini, his next relative Parelasmotherium represents, with which it forms a clade and assigned due to the location of the horn on the nasal bone of the sub-tribe of Iranotheriina. It faces that of Sinotherium and Elasmotherium, the Elasmotheriina.
Some researchers also provide Ningxiatherium directly to Parelasmotherium,but both show significant differences. Thus, Ningxiatherium has a partially ossified nasal bone, which does not occur in Parelasmotherium. Also, the occipital bone is strongly pulled out and pointed in the former, while it has a short and rectangular shape in the latter.
Ningxiatherium developed in the late Miocene about 11 million years ago, probably from Parelasmotherium, which had survived as the only representative of the Elasmotheriini in northern China after a cooler climatic phase, due to the extinction of numerous rhino species. How long it had lived is unclear, but it has not been proven in the Pliocene.
The first description was made by Chen Guafeng in 1977 based on the skull found in the Ningxia Basin. Today, two types are recognized:
Ningxiatherium longirhinus Chen, 1977
Ningxiatherium euryrhinus Deng, 2008

Iranotherium was a genus of rhinoceros that was once fairly widespread across central Asia. Individuals of Iranotherium were between three and four meters long,with an estimated mass of 3.6–4 tonnes,though as with other known kinds of rhinoceroses, the males would have been larger than females. The largest males could approach sizes up to four meters long. The disappearance of Iranotherium coincides with the rise of the genus Sinotherium, indicating that Sinotherium may have evolved from Iranotherium, and/or outcompeted the earlier genus. The Sinotherium genus is believed to eventually give rise to the better known Elasmotherium.
Sinotherium lagrelii

Sinotherium is a genus of extinct rhinoceros that seems to have been broadly spread across Eurasia between what is now Russia and northern China. The Sinotherium genus is has been believed to have evolved from the earlier Iranotherium. In turn Sinotherium is believed to have given rise to the much better known Elasmotherium.
Sinotherium was a large rhinoceros much heavier (up to 7 tonnes) than the largest modern African white rhino (3.2.6 tonnes). If it lived by the river it would have been easy for it to get stuck in the wet mud. More likely, S. lagrelii lived in an open, usually dry environment in northern China where droughts frequently occurred (Figure 3). Early East Asian large elasmothere populations may have frequented steppe environments more often than their wet-adapted descendants in southern Russia. S. lagrelii is phylogenetically near the root of the frontal-horned elasmothere radiation. Therefore, reconstructing the species as inhabiting the steppes is an alternative to the proposal that the frontal-horned elasmotheres lived in wetter habitats near rivers.
The giant woolly rhino Elasmotherium reached 20 ft long and 6.6 ft high.

Elasmotherium ("Thin Plate Beast") is an extinct genus of giant rhinoceros endemic to Eurasia during the Late Pliocene through the Pleistocene, documented from 2.6 Ma to as late as 50,000 years ago, possibly later, in the Late Pleistocene, an approximate span of slightly less than 2.6 million years. Three species are recognised. The best known, E. sibiricum, was the size of a mammoth and is thought to have borne a large, thick horn on its forehead this horn was used for defense, attracting mates, driving away competitors, sweeping snow from the grass in winter and digging for water and plant roots. Like all rhinoceroses, elasmotheres were herbivorous. Unlike any others, its high-crowned molars were ever-growing. Its legs were longer than those of other rhinos and were adapted for galloping, giving it a horse-like gait.
The fossil received its name from Johann Fischer von Waldheim,the Dirйcteur Perpйtuel of the Natural History Museum, Moscow University, at a presentation before the Societй Impйriale des Naturalistes in 1808. The next year in the Mйmoires of the society he reported what he had said in the Programme d"invitation:
Elasmotherium is an animal with an elongated head without incisors or canines, with 5 molars on each side [made] from sinuous layers.Then he noted on his derivation of the name:
From the Greek word ἐλασμος [elasmos], layer, to designate the layered form of the molar teeth.
All he had before him was one lower jaw donated to the museum by Yekaterina Romanovna Vorontsova-Dashkova, which he named Elasmotherium sibiricum, lamenting that it was the sole species of which he knew. The molars, the only teeth in the jaw, had formed in layers like tree rings, except the "rings", or lamellae, were highly corrugated. The edges in the grinding surface were elaborately sinuous to better break down the grasses on which the animal fed.
Various theories of Elasmothere morphology, nutrition and habits have been the cause of wide variation in reconstruction. Some show the beast trotting like a horse with a horn; others hunched over with head to the ground, like a bison, and still others immersed in swamps like a hippopotamus. The use of the horn and whether or not there was one, and how large, have been popular topics. The statistical correlations of modern paleontology have taken much of the speculation out of the subject, although some details remain undetermined.

The known specimens of E. sibiricum reach up to 4.5 m (15 ft) in body length with shoulder heights over 2 m (6 ft 7 in) while E. caucasicum reaches at least 5 m (16 ft) in body length with an estimated mass of 3.6–4.5 tonnes (4–5 short tons), based on isolated molars that significantly exceed those known from the Siberian species. Both species were among the largest in the family Rhinocerotidae, comparable in size to the woolly mammoth and larger than the contemporary woolly rhinoceros. The feet were unguligrade, the front larger than the rear, with 4 digits at the front and 3 at the rear.
Herbivores can be divided into two general groups on the basis of nutrition, which grade into each other morphologically: "foregut fermenters" and "hindgut fermenters". The border region is correlated to bulk: up to 600 to 1,200 kilograms (1,300 to 2,600 lb) are the former; over it, the latter. In foregut fermentation, the animal must "browse" to select the most nutritious plants and then ruminate to make up for the shorter digestive tract. The hind-gut fermenters are "bulk-feeders": they ingest large quantities of low-nutrient food, which they process for a longer time in a much longer intestine. The main food in that category is grass, indicating that Elasmotherium, like the elephants, was probably a grassland "grazer" moving over long distances to take advantage of the growth phases of grass in different regions. The standard is not without exception, as Indricotherium, the largest land mammal ever, with a weight of 15–20 tons, subsisted by browsing the treetops.
There are also paleontological indications of the grazing. In general, the normal position of the head can be determined by the angle between a vertical plane coinciding with the occiput of the cranium, which is always vertical, and a plane through the base of the cranium. A right or acute angle would indicate a head held high for browsing leaves at various heights. Elasmotherium had the most obtuse angle of the Rhinocerotids. It could only reach the lowest levels and therefore must have grazed habitually. This morphological feature favors the identification of the one-horned beast depicted in Rouffignac Cave (shown in this article) as Elasmotherium and lends some validity to the bison-like restorations based on it.
A third type of evidence that Elasmotherium was a grazer is that of tooth wear and morphology. Like all Rhinocerotidae, E. had cheek teeth evolved for herbivory: two premolars and three molars (originally taken for five molars), no incisors, no canines. Where some of the browsers kept the incisors in the form of tusks, E, had instead a spoon-like symphysis, or tip, of the lower mandible and a rostrum, or beak, of the upper, which served as a bony basis for a soft-tissue labial grasping and tearing mechanism. Grass, a very tough, fibrous material, contains phytoliths, microscopic granules mainly of silica, which act as sandpaper on the molars of grazers. Their response in geologic time is to evolve cheek teeth with large crowns (hypsodonty). There appears to be a correlation between grazing and hypsodonty.
As a general rule, extant herbivores with low-crowned teeth are predominantly browsers and species with high-crowned teeth are predominantly grazers.
Vladimir Onufryevich Kovalevsky first proposed a connection between hypsodonty and grazing for horses in 1873. Since then the concept has been expanded to all mammalian grazers at any time and has further been elaborated into hypsodonty or proto-hypsodonty and hypselodonty or euhypsodonty. The euhypsodonts, of which, surprisingly for its bulk, E. was one, have ever-growing high-crowned teeth. Most other examples are to be found among diminutive mammals such as Rodentia, which already casts doubt on the correlation, as they do not generally graze grass.
Teeth form from the top down through the deposition of enamel on a cement core by formative soft tissue in the jaw. The enamel of hypsodont Perissodactyla is highly rugose rather than sharp. In brachydont species, such as humans, when the crown is complete, the roots are deposited and finally the completed tooth erupts. Hypselodonty is a condition of tooth eruption and continued crown formation before a delayed root formation. In its most developed variety, the roots never form. Only rare fossils of E. show any sign of a root, and that on a premolar. No molars have roots, or, in the terminology of some, the roots are "open".
If all grazers are hypsodont, not all hypsodonts are grazers. The supposed correlation between grass-eating and hypsodonty proved difficult to support in a number of instances. Koenigswald, for example, pointed out that hypsodonty had occurred among the Gondwanatheria of the Mesozoic, a group of mammals so primitive that he describes their cheek teeth as "molariform", as they are neither clearly molars nor premolars.Molariform hypsodonty "cannot be correlated with a grass diet, since grasses were not present". Instead he suggests for his example, Sudamerica ameghinioi, that it lived a "semiaquatic and perhaps a burrowing way of life". Modern species provide many examples, from beavers to hippopotamuses.
Attempts have been made to link the wear on Elasmotherium teeth to grazing. In 1938, H.E. Wood, a Rhinocerotid tooth specialist, pointed out that interproximal wear, or loss of tooth surface between teeth, due to abrasion during mastication, of Elasmotherium is similar to that of the white rhinoceros, the only remaining Rhinocerotid grazer, which has hypsodont teeth. Data such as this led to an intuitive concept of some sort of correlation between grass-eating and hypsodonty, but it has been difficult to isolate mathematically.
A 2007 study by Mendoza and Palmqvist compared the habitats and diets of 134 species of living ungulates, for which this data is known, both Artiodactyla and Perissodactyla, against "32 craniodental measurements" to discover what correlations exist for the group studied and to test hypotheses concerning hypsodonty and mode of life. Habitats considered were open (savanna, deserts), mixed (wooded savanna, brush) and closed (riverine and forest). Diets considered were grazing, mixed grazing and browsing, browsing, omnivory and special niches, such as treetop browsing. Measurements included the Hypsodonty Index (HI) and Muzzle Width (MZW). The results showed that, except for the "high-level" browsers, hypsodonty is correlated to "open and mixed habitats". The HI was not precise enough to discriminate between open and mixed. However, high MZW is correlated to grazing in the open category, although some forest species also have wide muzzles. Grazers therefore are distinguished by a combination of high-crowned cheek teeth and wide muzzles, both of which are possessed by Elasmotherium. Life in the open is implied.
The preponderance of evidence is that Elasmotherium, as far as is known now, was a grassland grazer. It is in this context that Deng and Zheng, experts in the few surviving leg bones, conclude, concerning the morphology of the legs
Combined with hypsodont cheek teeth with much cement and strong enamel plication, the slender distal limb bones of E. caucasicum indicate that it is cursorial and dwells in an open steppe as a typical grazer.
By distal they mean "furthest outward"; that is, the extremities. Cursorial animals are unequivocally "runners" although the authors did not examine what sort of gait scientifically should be proposed as "running". They selected caucasicum for study because of the availability of a few dozen limb bone fragments from Nihewan, China. These made possible a selective comparison with the fewer bones remaining of other fossil rhinocerotids. In comparison with them, the long legs of Elasmotherium. are the most derived; that is, the others did not have the same cursorial capabilities. The authors approach but do not solve the problem of how to reconcile the weight with the supposed mobility. They say elsewhere in the article that the legs of caucasicum are to be distinguished from those of other fossil Rhinocerotids at Nihewan by their "enormous size".
The white rhinoceros at an estimated weight of 2.5 ton has been photographed galloping at a speed of about 30 km/h (19 mph). In a gallop, all feet are off the ground ("ballistic phase") twice a cycle, a feat that elephants, at 2.5-11 ton, cannot perform. They can walk up to 20 km/h (12 mph); however, their straight, relatively inflexible legs are those of striders, not the bent and spring-like legs of gallopers, which utilize haunches, ankle mobility and knee flexion to spring off the ground on alternative legs of a pair. Elasmotherium legs are sufficiently like those of the White Rhino to hypothesize a similar gait even though Elasmotherium weighed 4.5-5 ton.
The Ceratomorpha are so-called because their families, such as the Rhinocerotidae, of which Elasmotherium is indisputably one, are characterized by the presence of hooves, or horns and hooves, made of keratin, the same substance of which hair is made. These keratin structures appear to have formed in the Mesozoic, a remnant in humans being the nails. A keratin horn is to be distinguished from a bone horn and a tusk. Bone forms the base of most horns, but in some cases the horn is entirely of bone. A tusk is a modified canine or incisor tooth. Rhinocerotidae have had tusks, but not Elasmotherium. Two open questions are whether they were horned or hornless, hairy or hairless. Most Rhinocerotidae have and have had horns, but there are some instances of hornlessness, and most are or were hairy, such as the Wooly Rhinoceros, but no instances of hair or horn have yet been found for Elasmotherium. Only circumstantial evidence of them exists.
The main evidence suggestive of a horn on Elasmotherium is a frontal protuberance, which struck the attention of the late 19th century paleontologists and was immediately interpreted as the bony basis for a horn by most investigators from that time forward. A skull of E. sibiricum from the Volga region (cast shown in this article"s lead picture) described by Alexander Brandt in the Russian journal, Niwa, and reported in Nature in 1878 offers the following description of the protuberance: hemispherical, 5 inches (13 cm) deep, furrowed surface, circumference of 3 feet (0.91 m). The furrows are interpreted as the seats of blood vessels for the tissues that generated the horn.
The whole analogy with the rhinoceros points with the greatest certainty to the previous existence of a horn, which, to judge from the size of the blood vessels once encircling the base, must have possessed enormous dimensions.
Brandt was already familiar with the legend of a unicorn among the Tatars of Siberia with a horn so large it required a sledge for transport, and made the connection in interpreting the bump as the base of a horn. He also interpreted the rostrum of the upper mandible as the basis of a nasal horn, a hypothesis now rejected in favor of the cropping labia. In any case, the non-circular base indicates a section through the horn would not have been circular. This possibility is supported by another fossil with a non-circular partially healed puncture wound in the base, chiefly interpreted as the result of dueling other males with the horn.
The ungulates typically combine keratin and bone in various structures. If horns are keratinous, they have a bone core. Rhinocerotids horns, however, are uniquely derived.
This tissue is "strikingly convergent" with other "cornified epidermis" in horses, cetaceans, artiodactyls and birds. The horn is not attached to the bone of the boss but grows from the surface of a dense dermal tissue. The top layer keratinizes itself to form tubules about 1-2 millimeter high, the cells of which then die. The next layer forms below it. As the layers age the horn loses diameter by degradation of the keratin due to ultraviolet light, desiccation and mechanical wear from contact with objects and agonistic behavior. However, melanin and calcium deposits in the center harden the keratin there, causing differential wear and shaping of the horn.
The dermis generating the horn is anchored to the boss by interpenetration between rugosities – various irregularities of bone, which it creates by deposition. This tissue is a specialization of dermal armor, which, whenever it attaches to bone, deposits the rugosities to strengthen the attachment. The author states that rugosity is "a bony signature of dermal armor". Cranial rugosity is an indication, but not a sure sign, of a horn. If, on the other hand, an annular (ring-shaped) pattern is visible in the rugosity, it is due to "stress concentration at the edges of horns" and is "the signature for epidermal horns".
Hieronymus found annular rugosities in all living and some fossil Rhinocerotidae. The rings had previously been noted on additional fossils. To date Elasmotherium has not been examined for rings under lighting designed to show them up; however, based on the observations of other paleontologists, the author says "squamosal rugosity" is the "most pronounced cranial rugosity in the elasmotherine lineage" This fact suggests an especially firm attachment was required, which, combined with the extraordinarily large hump of muscle for managing the head, could suggest a large and heavy horn. In the early 19th century the state of the fossils had not yet revealed the presence of a horn. From around 1910 and ever since then, paleontologists have not ventured a mathematical estimate but rather have preferred occasionally to refer to the horn as immense, enormous, great or huge (these words turn up more often in popularizing works than in scientific articles). As the size and shape of the horn would depend on the concentration of melanin and calcium, and no known indicator of those remains, any further estimate of horn morphology is purely speculative.
The genus is known from hundreds of find sites, mainly of cranial fragments and teeth, but in some cases nearly complete skeletons of post-cranial bones, scattered over Eurasia from Eastern Europe to China. For example, Kazakhstan alone has 30 sites of E. sibiricum.Dozens of crania have been reconstructed and given archaeological identifiers. The division into species is based mainly on the fine distinctions of the teeth and jaws and the shape of the skull. The finds can be dated only by context.
Elasmotherium is believed to have descended from the Late Pliocene genus of Central Asia, Sinotherium. Elasmotherium is thought to be the most derived genus of elasmothere, with E. caucasicum in turn being more derived than E. sibiricum.The two Chinese fossils, formerly considered distinct species, E. inexpectatum and E. peii, defined by Chow in 1958, have been sunk into E. caucasicum. They were found in northern China from the Early Pleistocene Nihewan Faunal Assemblage (from the same valley as nearby Nihewan in Shanxi Province) and were extinct at approximately 1.6 Ma.
E. caucasicum, defined by Borissjak in 1914, flourished in the Black Sea region, as a member of the Early Pleistocene Tamanian Faunal Unit (1.1–0.8 Ma, Taman Peninsula). However, an elasmotherian species turned up in the preceding Khaprovian or Khaprov Faunal Complex, which was at first taken to be E. caucasicum, and then on the basis of the dentition was redefined as a new species, E. chaprovicum (Shvyreva, 2004), named after the Khaprov Faunal Complex. The Khaprov is in the MIddle Villafranchian, MN17, which spans the Piacenzian of the Late Pliocene and the Gelasian of the Early Pleistocene of Northern Caucasus, Moldova and Asia and has been dated to 2.6–2.2 Ma.
E. sibiricum, described by Johann Fischer von Waldheim in 1808 and chronologically the latest species of the sequence, coming from E. caucasicum in the Middle Pleistocene, ranged from southwestern Russia to western Siberia and southward into Ukraine and Moldova. It appears in the Middle Pleistocene Khazar Faunal Complex of the Sea of Azov region, which has "no exact stratigraphic situation".
The end of the elasmotheres is questionable; new evidence continues to turn up. The latest is from two caves in southern Siberia, a E. sibericum tooth in Smelovskaya Cave 11and remains from Batpak, both associated with Middle Pleistocene relict species, including large herbivores and predators. They must have been dragged into the caves by some predator, perhaps even modern man. The elasmothere tooth and one of a cave hyena from Smelovskaya were carbon dated to slightly greater than 50,000 BP in the Late Pleistocene.A recent study of relict Late Pleistocene remains in the Beringia region (Alaska, Eastern Siberia) identified a pattern of "imbedded micrometeorites" consistent with a space impact event similar to the Tunguska event of 1908, only carbon-dated to 37,000 BP. A Siberian Elasmotherium skull in a museum was found to have this pattern.
Тагове:
