

Прочетен: 4546 Коментари: 0 Гласове:
Последна промяна: 30.05.2020 17:42

Placoderms are arguably the most diverse groups of the early jawed fishes. They first appeared in the Early Silurian and diversified dramatically during the Devonian, but became extinct at the end of that period. About 200 genera of placoderms evolved during this interval, with the vast majority restricted to the Devonian. Some developed into the first vertebrate giants. Placoderms are armored by their extensive dermal skeleton. These dermal bones (or plates) form head and thoracic shields that are either articulated by distinctive joints or fused into a single unit. Pectoral fins are typically well developed. Bony shearing or crushing structures on the jaws substitute for true teeth, which are absent. The jaw joint is simple. Most placoderms are known only from plates of the head or thoracic shields, but some exceptionally preserved specimens reveal heterocercal (shark-like) tails, a pair of pelvic fins, and single dorsal fins; there is no clear evidence for anal fins.
The earliest identifiable placoderm fossils are from China and date to the early Silurian. At that time, they were already differentiated into antiarchs and arthrodires, as well as other, more primitive, groups. Earlier fossils of basal Placodermi have not yet been discovered.

The Silurian fossil record of the placoderms is both literally and figuratively fragmented. Until the discovery of Silurolepis (and then, the discoveries of Entelognathus and Qilinyu), Silurian-aged placoderm specimens consisted of fragments. Some of them have been tentatively identified as antiarch or arthrodire due to histological similarities; and many of them have not yet been formally described or even named. The most commonly cited example of a Silurian placoderm, Wangolepis of Silurian China and possibly Vietnam, is known only from a few fragments that currently defy attempts to place them in any of the recognized placoderm orders. So far, only three officially described Silurian placoderms are known from more than scraps:
1.The basal antiarch Silurolepis, from the Ludlow epoch of Yunnan, China, known from an almost complete thoracic armor.
2.Entelognathus, a placoderm incertae sedis that combines features of primitive arthrodires with jaw anatomy otherwise only seen in bony fish and tetrapods.
3.Qilinyu, a close relative of Entelognathus that further links Entelognathus as a transitional form between placoderms and other stem-gnathostomes and crown-group gnathostomes.
The first officially described and oldest Silurian placoderm is an antiarch, Shimenolepis, which is known from distinctively ornamented plates from the late Llandovery of Hunan, China. Shimenolepis plates are very similar to the early Devonian yunnanolepid Zhanjilepis, also known from distinctively ornamented plates.
Paleontologists and placoderm specialists suspect that the scarcity of placoderms in the Silurian fossil record is due to placoderms" living in environments unconducive to fossil preservation, rather than a genuine scarcity. This hypothesis helps to explain the placoderms" seemingly instantaneous appearance and diversity at the very beginning of the Devonian.
During the Devonian, placoderms went on to inhabit and dominate almost all known aquatic ecosystems, both freshwater and saltwater.But this diversity ultimately suffered many casualties during the extinction event at the Frasnian–Famennian boundary, the Late Devonian extinctions.The remaining species then died out during the Devonian/Carboniferous extinction event; not a single placoderm species has been confirmed to have survived into the Carboniferous.
Other than the earliest placoderms which come from the Early Silurian of China, the fossil record of placoderms is limited to the Devonian. All the major groups were present at the beginning of the Devonian so it’s apparent that a degree of diversification occurred sometime in the Middle and Late Silurian. Placoderms lost 6 out of 14 Late Frasnian families during the Frasnian/Famennian Mass Extinction, an event which is estimated to have eliminated 20% of all marine animal families. Most of the remaining families persisted until the very end of the Devonian. This second extinction event also claimed some early sharks, ray-fins and lobe-fin fishes, but apparently had a modest effect on other fauna.

There are six major groups of Devonian placoderms. One group,the Arthrodires, account for nearly 2/3 of the genera.
Arthrodiriformes

1. Aleosteus 2. Atlantidosteus 3. Coccosteus 4. Dunkleosteus 5. Dinichthys
Extinct order of armored placoderm fish with a joint between the armor surrounding the head and body and a mouth structure allowing for a relatively large mouth opening which lived from the Late Silurian to the late Devonian.
Arthrodires are distinguished by having two pairs of gnathal (tooth-like) plates extending from the upper jaw. The more advanced arthrodires also have an unusual neck joint that considerably increases gape size. Most placoderms were benthic but several of the more advanced arthrodires have reduced armor and were probably adept swimmers. Arthrodires are typically less than 1 m in length, but some (e.g., Dunkleostus) became the giants of their day.Coccosteus, Eastmanoesteus, Holonema and Rolfosteus are well known arthrodires from elsewhere.
Authorities differ on whether phyllolepids belong within the arthrodires or in a separate group. These dorsoventrally flattened fishes lack the second pair of gnathal plates and jointed neck that characterize most arthrodires, but they share some other features that may warrant inclusion.
Antiarchiformes

1. Pterichthyodes 2. Bothriolepis 3. Yunnanolepis 4. Pterichthys 5. Asterolepis
Extinct order of heavily armored placoderm fish with squared, box-shaped heads which lived from the early Silurian to the late Devonian.
Antiarchs are the second most diverse group of placoderms. They are also the strangest. The jointed head and thoracic shields formed a heavily armored box with a small anterior opening for two protruding eyes. Each pectoral fin was completely encased in bone and in the more advanced genera it was jointed in a fashion more reminiscent of arthropods than of vertebrates. Asterolepis, Bothriolepis and Remigolepis are antiarchs. An interesting side story concerns one antiarch, Bothriolepis. Serial sectioning of several well-preserved specimens by R.H. Dennison (1941) revealed a pair of sac-like structures that opened from the pharynx and extended well into the trunk. Dennison’s interpretation that these novel structures were lungs was not widely accepted. The case was taken up by Wells and Door (1985), who proposed that Bothriolepis could have been semi-terrestrial. They argued that the boxy armor supported the body out of water, the jointed pectorals helped the fish to move on land, and the sac-like structures allowed the fish to breath air.
Rhenaniformes
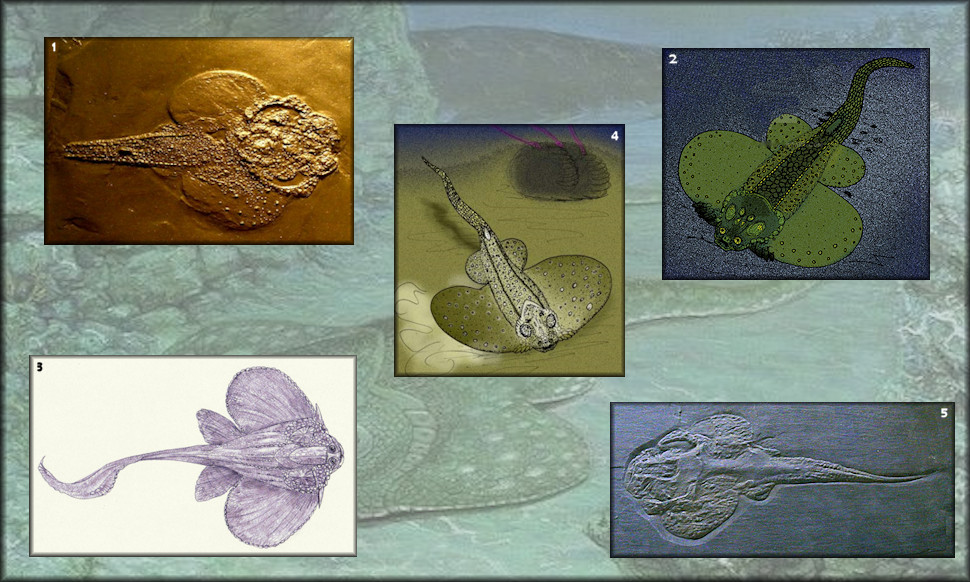
1. Ohioaspis 2. Bolivosteus 3. Jagorina 4. Asterosteus 5. Gemuendina
Extinct order of ray-like placoderm fish with armor made up of a mosaic of unfused scales and tubercles and lived from the early Devonian to the middle Devonian.
Rhenanids (e.g., Germendina) are even more dorsoventrally compressed placoderms with reduced thoracic armor and horizontally expanded pectoral fins that are reminiscent of skates and rays.
Ptyctodontiformes

1. Campbellodus 2. Rhamphodopsis Australoptyctodus 4. Materpiscis 5. Ctenurella
Extinct order of unarmored placoderm fish which superficially resemble modern day chimaeras (i.e., Holocephali) and lived from the early Devonian to the late Devonian.
Ptyctodontids (e.g., Cambellodus, Ctenurella and Rhamphodopis) have enlongate bodies, whip-like tails and reduced head and thoracic armor.
The males of one ptyctodontid, Rhamphodopis, have clasper-like intromittent organs similar to those found on chondrichthyans (sharks and their kin). This finding has been use by some authorities to propose that placoderms and chondrichthyans are related, but this view has not been widely accepted.
Petalichthyiformes

1. Lunaspis 2. Neopetalichthys 3. Notopetalichthys 4. Wijdeaspis 5. Diandongpetalichthys
Extinct order of relatively small armored placoderm fish with flattened bodies, splayed pectoral fins, exaggerated lateral spines, flattened bodies, and numerous tubercles that decorated all of the plates and scales of their armor which lived from the early Devonian to the late Devonian.
Peltalichyids (e.g., Lunaspis) are dorsoventrally compressed placoderms with shortened thoracic armor, expanded pectoral spines and dorsally oriented eyes.
Other Placoderms

1. Stensioella 2. Pseudopetalichthys 3. Paraplesiobatis 4. Tollichthys 5. Nessariostoma
Small Placoderm families (e.g., Stensioelliformes and Pseudopetalichthyiformes) along with genera of uncertain placement within the Placodermi class.
The largest placoderm was the 10 metres (33 ft) long Dunkleosteus.

Dunkleosteus is an extinct genus of arthrodire placoderm fish that existed during the Late Devonian period, about 380–360 million years ago. Some of the species, such as D. terrelli, D. marsaisi, and D. magnificus, are among the largest arthrodire placoderms ever to have lived.
The largest species, D. terrelli, measuring up to 10 m (33 ft) and weighing 3.6 t (4.0 short tons) was a hypercarnivorous apex predator. Few other placoderms, save, perhaps, its contemporary Titanichthys, rivaled Dunkleosteus in size.
Dunkleosteus is a pachyosteomorph arthrodire originally placed in the family Dinichthyidae, a family composed mostly of large, carnivorous arthrodires like Gorgonichthys. Anderson (2009) suggests, because of its primitive jaw structure, Dunkleosteus should be placed outside the family Dinichthyidae, perhaps close to the base of the clade Pachyosteomorpha, near Eastmanosteus. Carr and Hlavin (2010) resurrect Dunkleosteidae and place Dunkleosteus, Eastmanosteus, and a few other genera from Dinichthyidae within it.(Dinichthyidae, in turn, is made into a monospecific family).
New studies have revealed several features in both its food and biomechanics, as well as its ecology and physiology. Placodermi first appeared in the Silurian, and the group became extinct during the transition from the Devonian to the Carboniferous, leaving no descendants. The class persisted in the fossil record for at least 70 million years, in comparison to the 400-million-year-long history of sharks.
In recent decades, Dunkleosteus has achieved recognition in popular culture, with a large number of specimens on display, and notable appearances in entertainment media like Sea Monsters - A Walking with Dinosaurs Trilogy and River Monsters. Numerous fossils of some species have been found in North America, Poland, Belgium, and Morocco. The name Dunkleosteus combines the Greek osteus , meaning "bone", and Dunkle, in honor of David Dunkle of the Cleveland Museum of Natural History.
Due to its heavily armoured nature, Dunkleosteus was probably a relatively slow but powerful swimmer. It is thought to have dwelt in diverse zones of inshore waters. Fossilization tends to have preserved only the especially armoured frontal sections of specimens, thus it is uncertain what exactly the hind sections of this ancient fish were like. As such, the reconstructions of the hindquarters are often based on smaller arthrodires, such as Coccosteus, that had hind sections preserved. The most famous specimens of Dunkleosteus are displayed at the Cleveland Museum of Natural History. Others are displayed at the American Museum of Natural History and in the Queensland Museum in Brisbane, Queensland. Instead of teeth, Dunkleosteus possessed two pairs of sharp bony plates which formed a beak-like structure. Dunkleosteus, together with most other placoderms, may have also been among the first vertebrates to internalize egg fertilization, as seen in some modern sharks.
Dunkleosteus terrelli possessed a four bar linkage mechanism for jaw opening that incorporated connections between the skull, the thoracic shield, the lower jaw and the jaw muscles joined together by movable joints.This mechanism allowed D. terrelli to both achieve a high speed of jaw opening, opening their jaws in 20 milliseconds and completing the whole process in 50-60 milliseconds, comparable to modern fishes that use suction feeding to assist in prey capture and produce high bite forces when closing the jaw, estimated at 6,000 N (1,350 lbf) at the tip and 7,400 N (1,660 lbf) at the blade edge in the largest individuals.The pressures generated in those regions were high enough to puncture or cut through cuticle or dermal armor suggesting that D. terrelli was perfectly adapted to prey on free-swimming, armored prey like arthropods, ammonites, and other placoderms.Frequently, fossils of Dunkleosteus are found with boluses of fish bones, semidigested and partially eaten remains of other fish.As a result, the fossil record indicates it may have routinely regurgitated prey bones rather than digest them.
Morphological studies on the lower jaws of juveniles of D. terrelli reveal they were proportionally as robust as those of adults, indicating they already had the ability to produce high bite forces and likely were able to shear into resistant prey tissue similar to adults, albeit on a smaller scale. This pattern is in direct contrast to the condition common in tetrapods in which the jaws of juveniles are more gracile than in adults.
At least 10 different species of Dunkleosteus have been described so far.
The type species, D. terrelli, is the largest, best-known species of the genus. It has a rounded snout. D. terrelli"s fossil remains are found in Upper Frasnian to Upper Famennian Late Devonian strata of the United States (Huron and Cleveland Shales of Ohio, the Conneaut of Pennsylvania, Chattanooga Shale of Tennessee, Lost Burro Formation, California, and possibly Ives breccia of Texas) and Europe.
D. belgicus (?) is known from fragments described from the Famennian of Belgium. The median dorsal plate is characteristic of the genus, but, a plate that was described as a suborbital is apparently an anteriolateral plate.
D. denisoni is known from a small median dorsal plate, typical in appearance for Dunkleosteus, but much smaller than normal.
D. marsaisi refers to the Dunkleosteus fossils from the Lower Famennian Late Devonian strata of the Atlas Mountains in Morocco. It differs in size, the known skulls averaging a length of 35 cm and in form to D. terrelli. In D. marsaisi, the snout is narrower, and a postpineal fenestra may be present. Many researchers and authorities consider it a synonym of D. terrelli.H. Schultze regards D. marsaisi as a member of Eastmanosteus.
D. magnificus is a large placoderm from the Frasnian Rhinestreet Shale of New York. It was originally described as "Dinichthys magnificus" by Hussakof and Bryant in 1919, then as "Dinichthys mirabilis" by Heintz in 1932. Dunkle and Lane moved it to Dunkleosteus in 1971.
D. missouriensis is known from fragments from Frasnian Missouri. Dunkle and Lane regard them as being very similar to D. terrelli.
D. newberryi is known primarily from a 28-cm-long infragnathal with a prominent anterior cusp, found in the Frasnian portion of the Genesee group of New York, and originally described as "Dinichthys newberryi".
D. amblyodoratus is known from some fragmentary remains from Late Devonian strata of Kettle Point, Canada. The species name means "blunt spear" and refers to the way the nuchal and paranuchal plates in the back of the head form the shape of a blunted spearhead. Although it is known only from fragments, it is estimated to have been about 20 ft long in life.
D. raveri is a small, possibly 1-m-long species known from an uncrushed skull roof, found in a carbonate concretion from near the bottom of the Huron Shale, of the Famennian Ohio Shale strata. Besides its small size, it had comparatively large eyes. Because D. raveri was found in the strata directly below the strata where the remains of D. terrelli are found, D. raveri may have given rise to D. terrelli. The species name commemorates Clarence Raver of Wakeman, Ohio, who discovered the concretion where the holotype was found.
Тагове:
Encyclopedia Largest prehistoric animals...
Encyclopedia Largest prehistoric animals...
