

Прочетен: 6064 Коментари: 0 Гласове:
Последна промяна: 29.04.2019 20:54
Turtles and tortoises (Testudines)

Turtles and tortoises comprise Order Testudines, evolutionarily one of the oldest groups of reptiles, which pre-dates the dinosaurs. The most prominent unifying characteristic of turtles is the presence of a shell, which evolved from an expansion of the ribcage. Turtle shells can be highly variable in size, shape and feel; some cartilagenous shells appear leathery, while others are hardened by the presence of horny scales called scutes. All turtles have a rigid beak in lieu of teeth, the shape of which is adapted to their food source. Since turtles are found in almost every type of habitat, species have developed specific adaptations with regard to their front legs and feet. Terrestrial turtles (like tortoises) have small, stumpy feet. Aquatic species possess webbed feet with claws for climbing onto logs and banks. Marine species use flippers to navigate through the water.
As cold-blooded organisms, turtles regulate their internal temperature through behaviors, such as basking in the sun. All turtles lay leathery eggs in nests created in the dirt or sand near their habitat. For some species, the sex of the turtles is determined by the temperature at which the eggs are incubated. Regardless of whether they live in an aquatic or terrestrial habitat, all turtles obtain their oxygen from the air.

Based on body fossils, the first proto-turtles are believed to have existed in the late Triassic Period of the Mesozoic era, about 220 million years ago, and their shell, which has remained a remarkably stable body plan, is thought to have evolved from bony extensions of their backbones and broad ribs that expanded and grew together to form a complete shell that offered protection at every stage of its evolution, even when the bony component of the shell was not complete. This is supported by fossils of the freshwater Odontochelys semitestacea or "half-shelled turtle with teeth", from the late Triassic, which have been found near Guangling in southwest China. Odontochelys displays a complete bony plastron and an incomplete carapace, similar to an early stage of turtle embryonic development.Prior to this discovery, the earliest-known fossil turtle ancestors, like Proganochelys, were terrestrial and had a complete shell, offering no clue to the evolution of this remarkable anatomical feature. By the late Jurassic, turtles had radiated widely, and their fossil history becomes easier to read.

Their exact ancestry has been disputed. It was believed they are the only surviving branch of the ancient evolutionary grade Anapsida, which includes groups such as procolophonids, millerettids, protorothyrids, and pareiasaurs. All anapsid skulls lack a temporal opening while all other extant amniotes have temporal openings (although in mammals, the hole has become the zygomatic arch). The millerettids, protorothyrids, and pareiasaurs became extinct in the late Permian period and the procolophonoids during the Triassic.
However, it was later suggested that the anapsid-like turtle skull may be due to reversion rather than to anapsid descent. More recent morphological phylogenetic studies with this in mind placed turtles firmly within diapsids, slightly closer to Squamata than to Archosauria.All molecular studies have strongly upheld the placement of turtles within diapsids; some place turtles within Archosauria,or, more commonly, as a sister group to extant archosaurs,though an analysis conducted by Lyson et al. (2012) recovered turtles as the sister group of lepidosaurs instead.Reanalysis of prior phylogenies suggests that they classified turtles as anapsids both because they assumed this classification (most of them studying what sort of anapsid turtles are) and because they did not sample fossil and extant taxa broadly enough for constructing the cladogram. Testudines were suggested to have diverged from other diapsids between 200 and 279 million years ago, though the debate is far from settled.Even the traditional placement of turtles outside Diapsida cannot be ruled out at this point. A combined analysis of morphological and molecular data conducted by Lee (2001) found turtles to be anapsids (though a relationship with archosaurs couldn"t be statistically rejected).Similarly, a morphological study conducted by Lyson et al.. (2010) recovered them as anapsids most closely related to Eunotosaurus. A molecular analysis of 248 nuclear genes from 16 vertebrate taxa suggests that turtles are a sister group to birds and crocodiles (the Archosauria).The date of separation of turtles and birds and crocodiles was estimated to be 255 million years ago. The most recent common ancestor of living turtles, corresponding to the split between Pleurodira and Cryptodira, was estimated to have occurred around 157 million years ago.The oldest definitive crown-group turtle (member of the modern clade Testudines) is the species Caribemys oxfordiensis from the late Jurassic period (Oxfordian stage).Through utilizing the first genomic-scale phylogenetic analysis of ultraconserved elements (UCEs) to investigate the placement of turtles within reptiles, Crawford et al. (2012) also suggest that turtles are a sister group to birds and crocodiles (the Archosauria).

The first genome-wide phylogenetic analysis was completed by Wang et al. (2013). Using the draft genomes of Chelonia mydas and Pelodiscus sinensis, the team used the largest turtle data set to date in their analysis and concluded that turtles are likely a sister group of crocodilians and birds (Archosauria).This placement within the diapsids suggests that the turtle lineage lost diapsid skull characteristics as it now possesses an anapsid-like skull.
The earliest known fully shelled member of the turtle lineage is the late Triassic Proganochelys. This genus already possessed many advanced turtle traits, and thus probably indicates many millions of years of preceding turtle evolution; this is further supported by evidence from fossil tracks from the Early Triassic of the United States (Wyoming and Utah) and from the Middle Triassic of Germany, indicating that proto-turtles already existed as early as the Early Triassic. Proganochelys lacked the ability to pull its head into its shell, had a long neck, and had a long, spiked tail ending in a club. While this body form is similar to that of ankylosaurs, it resulted from convergent evolution. Turtles are divided into two extant suborders: Cryptodira and Pleurodira. The Cryptodira is the larger of the two groups and includes all the marine turtles, the terrestrial tortoises, and many of the freshwater turtles. The Pleurodira are sometimes known as the side-necked turtles, a reference to the way they retract their heads into their shells. This smaller group consists primarily of various freshwater turtles.
The largest sea turtle ever was Archelon ischyros at 4 m (13 ft) long, 4.9 m (16 ft) wide and 2200 kg (4,850 lbs). 
The first specimen of Archelon (YPM 3000) was collected from the Campanian-age Pierre Shale of South Dakota (a geological formation dated to 80.5 million years ago)by Dr. G.R. Wieland in 1895 and described by him the following year (Wieland, 1896). The largest Archelon fossil, found in the Pierre Shale of South Dakota in the 1970s, measures more than 4 metres (13 ft) long, and about 4.9 metres (16 ft) wide from flipper to flipper. It was a marine turtle, whose closest living relative in the present day is the leatherback sea turtle. Archelon lived at a time when a shallow sea covered most of central North America. Most of the known remains have been found in South Dakota and Wyoming. Though anatomically similar to the earlier species Protostega gigas, it was much larger.
Unlike most modern turtles, Archelon did not have a solid shell, but instead had a skeletal framework supporting a leathery or bony carapace. Other distinguishing features include a pointed tail, a narrow skull, a relatively narrow, high-vaulted shell, and a pronounced overbite.The live weight of an Archelon ischyros is estimated at more than 2200 kg (4,850 lb). They probably had a very strong bite, and were optimized for feeding on pelagic mollusks such as squid.The specimen exhibited by the Museum of Natural History in Vienna is estimated to have lived to be a century old, and may have died while brumating on the ocean floor.
Although a turtle, Archelon has become a staple species that appears in almost every book about dinosaurs and other prehistoric animals. This almost mandatory inclusion comes from the fact that Archelon is the largest turtle ever known to exist and it lived at the end of the Cretaceous. For the most part dinosaurs were probably not a threat, as assuming Archelon laid eggs like turtles do today, it would only ever venture onto land to lay eggs. Conversely however these eggs would eventually hatch and the young may have had to run a gauntlet of predators on their way to the ocean. Back in the Cretaceous this could mean everything from small beach combing dinosaurs, primitive seabirds similar to Ichthyornis to even pterosaurs.
Archelon is usually envisioned as being very similar to the leatherback turtle, and likely had a similar preference for eating jellyfish and cephalopods. The horned beak of the mouth which had a clear overbite would have been very effective at snipping soft bodied animals like these into bite sized portions that could be swallowed. Another similarity between these two turtles is that neither one has a solid shell. Instead a series of bony struts, that in Archelon are actually the ribs, create a framework of bone that an outer and relatively thin carapace sits on top of.

Perhaps the easiest explanation for Archelon’s shell design is that of neutral buoyancy. This is where a marine animal’s body adapts so that it neither floats to the surface, nor sinks to the bottom. In order to achieve this effect the body changes along such lines as denser thick bones that counter the lifting effect of the air in the lungs so that the animal does not bob around on the surface. Good examples for marine adaptation in reptiles can be seen in the placodonts from the Triassic which were actually negatively buoyant so that they could sink to the bottom where their preferred food was.
Archelon however was not a bottom feeder, it had to stay active in the mid to upper surface areas where its prey was most abundant, and as such it needed to be neutrally buoyant so that it could adapt to different depths wherever its prey was. As a large turtle Archelon obviously had a large shell, but if this shell was solid bone with a horn covering like in smaller turtles it would be extremely heavy. It’s feasible that a heavy solid shell would have tipped the scale into negative buoyancy so that Archelon would have had to spend a greater amount of energy just to stay up in the areas its food supply was, as well as rising up to breathe. This would not suit any animal, especially large ones like Archelon.
A shell composed of a framework with a relatively thin covering however would actually be very lightweight as well as reducing the body density so that Archelon did not readily sink as well as a more solid creature. This in turn would allow Archelon to cruise the oceans at a more leisurely pace and conserve energy so that it would not need as high a calorie intake as it would have done if the shell were solid.
The presence of large mosasaurs such as Tylosaurus as well as possibly even sharks like Cretoxyrhina suggest that Archelon probably was not invulnerable to predators, particular from attacks to the flippers. Despite this the sheer physical size of the shell may have still been enough to prevent some predators from being able to close their jaws around the body. At the very least this would have made a fully grown Archelon a difficult prey item compared to other softer bodied marine reptiles. Aside from this large size four star-shaped plates are found on the underside of the shell which seems to reinforce it. These may appear on the underside as a form of additional defence from predators that hit it from below. It’s perhaps not implausible that such attacks may have been initiated by mistake by predators that confused Archelon for a different kind of animal, possibly explaining the presence of these plates only on the underside.
The next largest was Protostega at 3 m (9.8 ft).

Protostega ("first roof") is an extinct genus of marine turtle containing a single species, Protostega gigas. Its fossil remains have been found in the Smoky Hill Chalk formation of western Kansas (Hesperornis zone, dated to 83.5 million years ago) and time-equivalent beds of the Mooreville Chalk Formation of Alabama. Fossil specimens of this species were first collected in 1871, and named by Edward Drinker Cope in 1872. With a length of 3 metres (9.8 ft), it is the second-largest sea turtle that ever lived, second only to the giant Archelon, and the third-largest turtle of all time behind Archelon and Stupendemys. Growing more than 10 feet (3 meters) long, Protostega was among the largest turtles to ever live. Unlike most turtles, whose shells are made of expanded and fused bones that form a relatively solid dome, widely spaced bones that looked more like the rafters of a roof held up Protostega"s leathery shell. Though the shell design provided less protection, the lighter load combined with powerful, flipper-like front legs made Protostega a strong, inexhaustible swimmer. Females likely migrated hundreds of miles to lay eggs on sandy beaches, much like sea turtles do today.
Movement onshore, however, was difficult. An adult female Protostega may have weighed a ton or more, a hefty load to drag out of the ocean to lay eggs. But lay eggs on the beach they did by the dozens, a reproductive strategy of safety in numbers that helped at least a few survive to adulthood. In fact, marine turtles were the only seagoing reptiles to escape extinction at the end of the Cretaceous, 65 million years ago.
A large and pointed head with a sharp beak and strong jaws probably helped the ancient turtles feed on slow-moving marine creatures such as jellyfish and shellfish as well as seaweed and floating carcasses—much as their descendents do today. Shark teeth embedded in Protostega bones housed at a museum in Chicago suggest the turtles were sometimes a meal themselves.
Atlantochelys
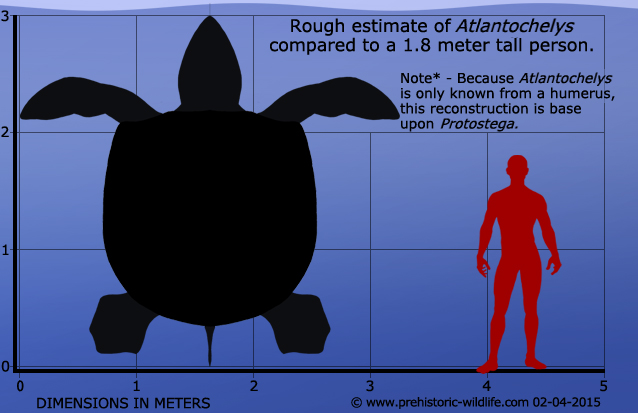
Atlantochelys was first named in 1849 and based upon the description of half a humerus (analogous to your upper arm bone). For well over one hundred and fifty years that was it, but then in 2012 the missing half of this bone was actually matched up to the original specimen. Although Atlantochelys may be considered dubious because of still a lack of overall fossil remains, the humerus of Atlantochelys is still identified as coming from a protostegid turtle, while at the same time not like other known specimens. Scaling the size of the humerus to relative genera has yielded an approximate estimate that Atlantochelys grew to about three meters in length. This large size would have been the best protection for Atlantochelys given that large marine reptiles called mosasaurs as well as large sharks with teeth especially adapted for cutting bone and shell like those of Cretoxyrhina were all swimming in the oceans at the same time as Atlantochelys.
Atlantochelys has been classed within the Protostegidae group of turtles, and so speculated reconstructions of Atlantochelys are usually based upon other members of this group such as Protostega and Archelon, both of which seem to have been larger than Atlantochelys.
The newly discovered turtle, named Ocepechelon bouyai, is one of the biggest marine turtles that ever lived on Earth.

The generic name Ocepechelon derives from an acronym for the Groupe Office Chйrifien des Phosphates, the mining company exploiting phosphatic deposits in Morocco, and from the Greek word chelone (turtle).
Ocepechelon is known from an almost complete 28-inch-long (70 cm) skull found in Sidi Chennane area in Morocco’s Khouribga Province.
Its striking anatomy suggests the turtle was adapted for suction feeding, never seen in known turtles.
Ocepechelon shares interesting resemblances with beaked whales. “Ocepechelon and most beaked whales are large animals, and, as in all other extant cetaceans and Ocepechelon, the nostrils of beaked whales are posterodorsally shifted. They also share similarities on their feeding apparatus: elongated jaws and a small gape.
The mouth gape of Ocepechelon can be inferred from the diameter of the tube – about 2.5 inches (6 cm). It is worth emphasizing the disproportion between the tiny size of the mouth opening and the large size of the Ocepechelon’s skull.
The snout diameter shows that Ocepechelon was a small-prey hunter and may have fed on small fishes, cephalopods and jellyfishes.
Тагове:
Encyclopedia Largest Prehistoric Animals...
Encyclopedia Largest prehistoric animals...
