

Прочетен: 7004 Коментари: 0 Гласове:
Последна промяна: 02.06.2019 19:45
Allosauridae

Allosauridae was a family of medium to large sized carnivorous theropod dinosaurs. Members included Saurophaganax, Allosaurus, and the poorly known Epanterias, which could well be a specimen of Allosaurus. Allosauridae was named by American paleontologist Othniel Charles Marsh in 1878.
The exact number of genera that can be included here is very debatable and most of the upper Jurassic and Lower Cretaceous carnosaurs are somewhat closely related to Allosaurus. Whatever the case, allosaurids seemed to be the most successful hunters of their times, outnumbering the Megalosauridae and Ceratosauridae, with which they competed for prey. The allosaurids would eventually be succeeded by their close relatives the carcharodontosaurs in the southern hemisphere, and replaced by the coelurosaurian tyrannosaurids in the northern hemisphere, during the Cretaceous Period. .png)
At one time the family Allosauridae included such diverse taxa as Allosaurus, Neovenator, and Acrocanthosaurus, and even all of the Carcharodontosaurs, as well as the poorly known Valdoraptor, Wakinosaurus and "Chilantaisaurus maortuensis." Nowadays everything apart from Allosaurus (and probably synonyms such as Saurophaganax) are placed in other families, specifically the Carcharodontosauridae. Unless giant forms such as Epanterias et al are given their own generic name, Allosaurus would have to revert to its senior synonym of Antrodemus, because there would then only be one genus of large Morrison carnosaur, and Antrodemus would no longer be indeterminate (such are the rules of scientific nomenclature). This would be the same situation as Triceratops and Torosaurus being renamed Agathaumas should they all turn out to be part of a single growth series.
While usually - apart that is from the little known giant species like A. amplexus - not in the same size range as T. rex, Allosaurus was an iconic member of the late Jurasisc megafuana, a huge and powerful predator, and clearly a very ecologically successful animal, as it easily outnumbered contemporary megapredators such as Ceratosaurs and Torvosaurus. An adult allosaur combined a flightless bird-like body, physiology, and agility with the brain of a crocodile and the mass of a medium-sized rhino; which means that it would basically lunge at anything that moved and looked like lunch(which basically was any other animal large enough to notice). Camptosaurs, stegosaurs, medium-sized sauropods, even other theropods, there was probably nothing short of a thirty-tonne apatosaur it wouldn"t take on. With its powerful jaws and hind limbs, strong grasping forelimbs, and long counterbalancing tail, this impressive yet graceful animal very much typified the large preditor ecotype of the mid-Mesozoic.
We can be pretty sure that Allosaurus evolved from something like Sinraptor or one of his cousins, and replaced them as the top carnivores of the Kimmeridgian and Tithonian ages (the latest Jurassic period). The two are pretty similiar in size and appearance and no doubt lifestyle, and differ only in detail, however Allosaurus has some advanced minor features not found in early or more primitive texa, as revealed in details of the skeleton (doubtless there were physiological and soft tissue details as well).
Morphologically (in terms of the evolution of form) as well as stratigraphically (the sequence in deep time revealed by fossil remains), Allosaurus represents the non-missing link between the Middle Jurassic Sinraptorids and the Cretaceous Carcharodontosaurs. If sinraptorids and equivalent taxa, such as Lourinhanosaurus, constituted the first pulse of allosauroidean evolution, then Allosaurus (or Allosaurines) were the second, and the dual Cretaceous adaptive radiations of giant carcharodontosaurs and medium-sized megaraptorans the third and culminating one. Here as elsewhere, the fossil record reveals the evolutionary history of life on Earth as beautiful sequence of transformation of form, with one type evolving into or being replaced by another through the course of deep time.

While some more fastidious phylogenetists might not want to state hypotheses too literally, citing the incompleteness of the fossil record as a reason to avoid statements about transitional forms or identification of any known taxa as actual objective (rather than hypothetical and abstract) ancestors, or rejecting the premise that cladograms provide insight into evolution in deep time, we would rather avoid such phylopessimism and adopt a more concrete and optimistic approach. We affirm that stratigraphy and the fossil record provide a strong phylogenetic signal which helps reveal details ofthe evolutionary tree of life.
This raises the problem of how such dendrograms should be drawn. Dendrograms such as the ones we employ here are not as strict or rigorous as cladograms, but they do provide popular representations of them (and are often confused with them). Cladistic methodology is - unlike evoluitionary systematics - not concerned with describing actual common ancestors; cladograms are simply abstract diagrams for conveying phylogenetic hypotheses, generally arrived at through statistical parsimony analysis. In such a diagram, Allosaurus might be shown on one terminal node, Sinraptor on another, and so on, without any comment as to whether one is the actual ancestor of the other. We say that Sinraptor is placed in a more basal position (as we have done here), but that doesn"t make it an ancestor in cladistic formalism.
Formalism can lead to problems though. When popular adaptions of such cladograms are drawn online (i.e. dendrograms), it may seem that all the species assigned to a clade appear as a distinct and quite dramatic evolutionary radiation, as in Mikko"s Phylogeny, Wikipedia dendrograms, or the Theropod Database, when all that is being done is representing sister-group relationships in simple diagramatic format. (scroll down for Allosaurus on the Theropod Database link). This is where Pattern Cladists get it right; such hypotheses should not be intended as literal geneologies. The problem is, at this poor level of resolution, it is often very difficult to know which species gave rise to which. So they tend to bunch into artificially monophyletic clusters (we saw this with groups like Ceratosauria and Ceolophysoidea). And even if (using here the linnaean genus and species hierarchy) a genus (a group of species) is ancestral to a later group (i.e. one of the species, known or unknown, in that genus directly evolved by Darwinian means into subsequent species, genera, and/or clades) traditional cladistics does not allow this to be represented (because it is only concerned with sister relationships, not ancestors and descendents).
Now, since Allosaurus is the only large advanced avetheropod known from the latest Jurassic, and assuming the phylogenetic hypothesis Sinraptoridae + (Allosaurus + (Neovenator + (Carcharodontosauridae + Megaptora)) or similar (e.g. (Carcharodontosauridae + (Neovenator + Megaptora))), it would be a paraphyletic taxon ( grade).
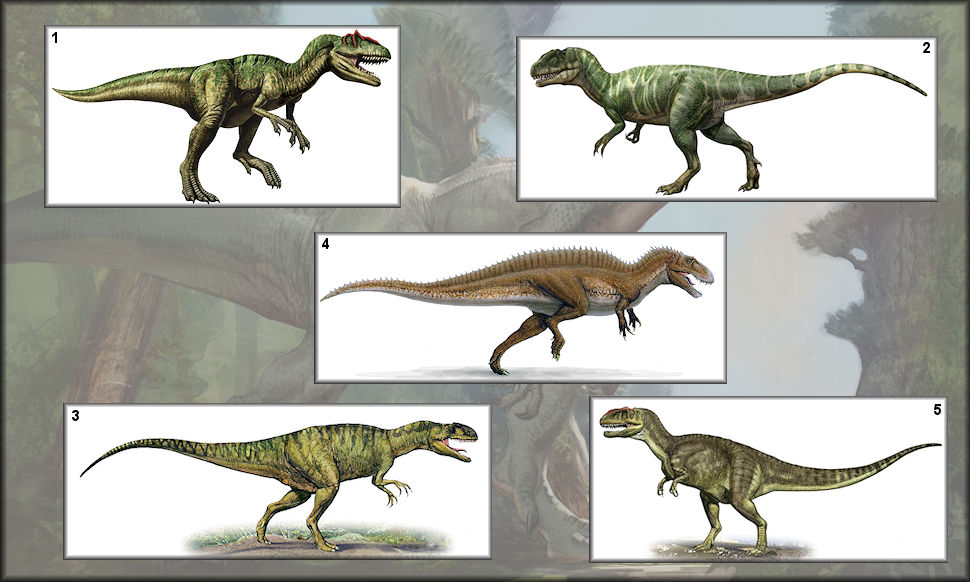
1. Allosaurus 2. Piatnitzkysaurus 3. Neovenator 4. Acrocanthosaurus 5. Yangchuanosaurus
Even if we don"t know which species of Allosaurus gave rise (via speciation) to the lineage culminating in early Cretaceous forms, it would still have been one of the species (or more than one, if they form a series), even if it is not one currently known to science, or even one that never fossilised.
The one on the left is a version of the type found everywhere online, which is based on a sort of cladistic literalism; as common ancestors are not referenced, we have instead a ghost lineage alongside a brief-flowering but ultimately sterile Allosaurus clade. The one on the right however is based on the premise that Allosaurus is a paraphyletic taxon, i.e. that it is the actual ancestor of Cretaceous allosauroidea. Of course, this still could be refuted if another advanced and intermediate late Jurassic taxon is found (in which case that taxon becomes the new transitional, paraphyletic grade), but on the basis of current knowledge. For this reason, we"re introducinga new method of drawing dendrograms. At least that goes for the present author, some of the time. Other Palaeos contributers will differ according to whether they are phylogenticists, evolutionary suystematists, evolutionary phylogenetists, stratocladists, or some other methodology or combination of methodologies. Hence there is unlikely to be any rigorous attempt at consistency in these webpages for quite some time, if at all.
Saurophaganax

Saurophaganax ("lizard-eater") is a genus of allosauriddinosaur from the Morrison Formation of Late JurassicOklahoma (latest Kimmeridgian age, about 151 million years ago), USA.Some paleontologists consider it to be a species of Allosaurus (A. maximus). Saurophaganax represents a very large Morrison allosaurid characterized by horizontal laminae at the bases of the dorsal neural spines above the transverse processes, and "meat-chopper" chevrons.The maximum size of S. maximus has been estimated at anywhere from 10.5 metres (34 ft)to 13 m (43 ft) in length and around 3 tonnes (3.0 long tons; 3.3 short tons) in weight.
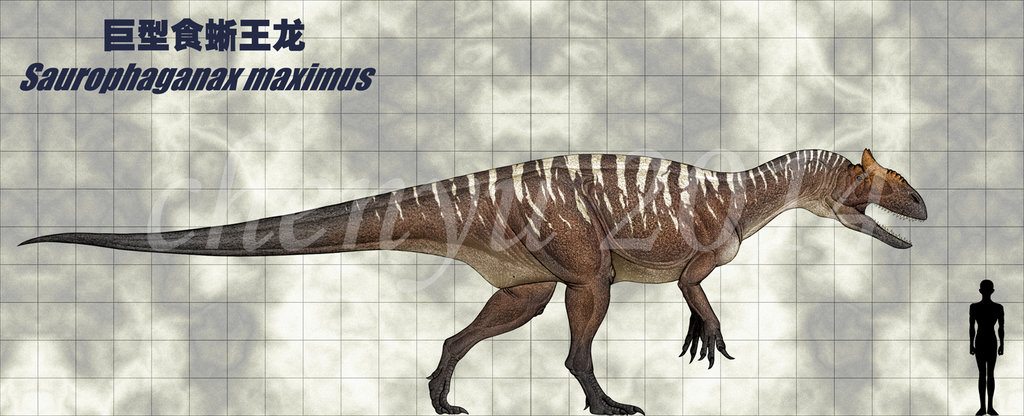
In 1931 and 1932 John Willis Stovall uncovered remains of a large theropod near Kenton in Cimarron County, Oklahoma in layers of the late Kimmeridgian. In 1941 these were named Saurophagus maximus by Stovall in an article by journalist Grace Ernestine Ray.The generic name is derived from Greek σαυρος, sauros, "lizard" and φάγειν, phagein, "to eat", with the compound meaning of "eater of saurians". The specific epithet maximus means "the largest" in Latin. Because the naming article did not contain a description, the name remained a nomen nudum. In 1950 Stovall described the finds.However, in 1987 Spencer George Lucas e.a. concluded a lectotype had to be designated among the many bones: OMNH 4666, a tibia.
Later it was discovered that the name Saurophagus was preoccupied: it had already in 1831 been given by William Swainson to a tyrant-flycatcher, a real eater of lizards.In 1995 Daniel Chure named a new genus: Saurophaganax, adding Greek suffix -άναξ, anax, meaning "ruler", to the earlier name. Chure also established that the lectotype tibia was not diagnostic in relation to Allosaurus. He designated another element as the type specimen: OMNH 01123, a neural arch. This was not intended as a neotype of the old genus but as the holotype of a genus different from "Saurophagus". The type species Saurophaganax maximus, based on diagnostic material, is thus not to be considered conspecific with Saurophagus maximus based on an undiagnostic bone — which species Chure later stated to be a nomen dubium — and Saurophaganax is not a renaming of "Saurophagus".Much of the material previously referred to Saurophagus maximus, namely those diagnostic elements that could be distinguished from Allosaurus, were by Chure referred to Saurophaganax maximus. They contain disarticulated bones of at least four individuals.
Saurophaganax is the official state fossil of Oklahoma and a large skeleton of Saurophaganax can be seen in the Jurassic hall in the Sam Noble Oklahoma Museum of Natural History. Although the best known Saurophaganax material was found in the panhandle of Oklahoma, possible Saurophaganax material, NMMNH P-26083, a partial skeleton including a femur, several tail vertebrae, and a hip bone, has been found in northern New Mexico.
The identification of Saurophaganax is a matter of dispute. It has been described as its own genus or as a species of Allosaurus: Allosaurus maximus.The most recent review of basal tetanurans accepted Saurophaganax as a distinct genus.New possible Saurophaganax material from New Mexico may clear up the status of the genus.
Saurophaganax was one of the largest carnivores of Late Jurassic North America. Chure even gave an estimate of 14 m (46 ft),though other estimations have been lower. The fossils known of Saurophaganax (both the possible New Mexican material and the Oklahoma material) are known from the Brushy Basin Member, which is the latest part of the Morrison Formation, suggesting that this genus was either always uncommon or that it first appeared rather late in the Jurassic. Saurophaganax was large for an allosaurid, and bigger than both its contemporaries Torvosaurus tanneri and Allosaurus fragilis. Being much rarer than its contemporaries, making up one percent or less of the Morrison theropod fauna, not much about its behavior is known.
The Morrison Formation is a sequence of shallow marine and alluvial sediments which, according to radiometric dating, ranges between 156.3 million years old (Ma) at its base, to 146.8 million years old at the top,which places it in the late Oxfordian, Kimmeridgian, and early Tithonianstages of the Late Jurassic period. This formation is interpreted as a semiarid environment with distinct wet and dry seasons. The Morrison Basin where dinosaurs lived, stretched from New Mexico to Alberta and Saskatchewan, and was formed when the precursors to the Front Range of the Rocky Mountains started pushing up to the west. The deposits from their east-facing drainage basins were carried by streams and rivers and deposited in swampy lowlands, lakes, river channels and floodplains.This formation is similar in age to the Solnhofen Limestone Formation in Germany and the Tendaguru Formation in Tanzania. In 1877 this formation became the center of the Bone Wars, a fossil-collecting rivalry between early paleontologists Othniel Charles Marsh and Edward Drinker Cope.
The Morrison Formation records an environment and time dominated by gigantic sauropod dinosaurs such as Barosaurus, Brontosaurus, Camarasaurus, Diplodocus, and Brachiosaurus. Dinosaurs that lived alongside Saurophaganax, and may have served as prey, included the herbivorous ornithischians Camptosaurus, Dryosaurus, Stegosaurus, and Othnielosaurus. Predators in this paleoenvironment included the theropods Torvosaurus,Ceratosaurus, Marshosaurus, Stokesosaurus, Ornitholestes, and Allosaurus, which accounted for 70 to 75% of theropod specimens and was at the top trophic level of the Morrison food web.Other vertebrates that shared this paleoenvironment included ray-finned fishes, frogs, salamanders, turtles, sphenodonts, lizards, terrestrial and aquatic crocodylomorphans, and several species of pterosaur. Early mammals were present in this region, such as docodonts, multituberculates, symmetrodonts, and triconodonts. The flora of the period has been revealed by fossils of green algae, fungi, mosses, horsetails, cycads, ginkgoes, and several families of conifers. Vegetation varied from river-lining forests of tree ferns, and ferns (gallery forests), to fern savannas with occasional trees such as the Araucaria-like conifer Brachyphyllum.In Oklahoma, Stovall unearthed a considerable number of Apatosaurus specimens, which may have represented possible prey for a large theropod like Saurophaganax.
Epanterias
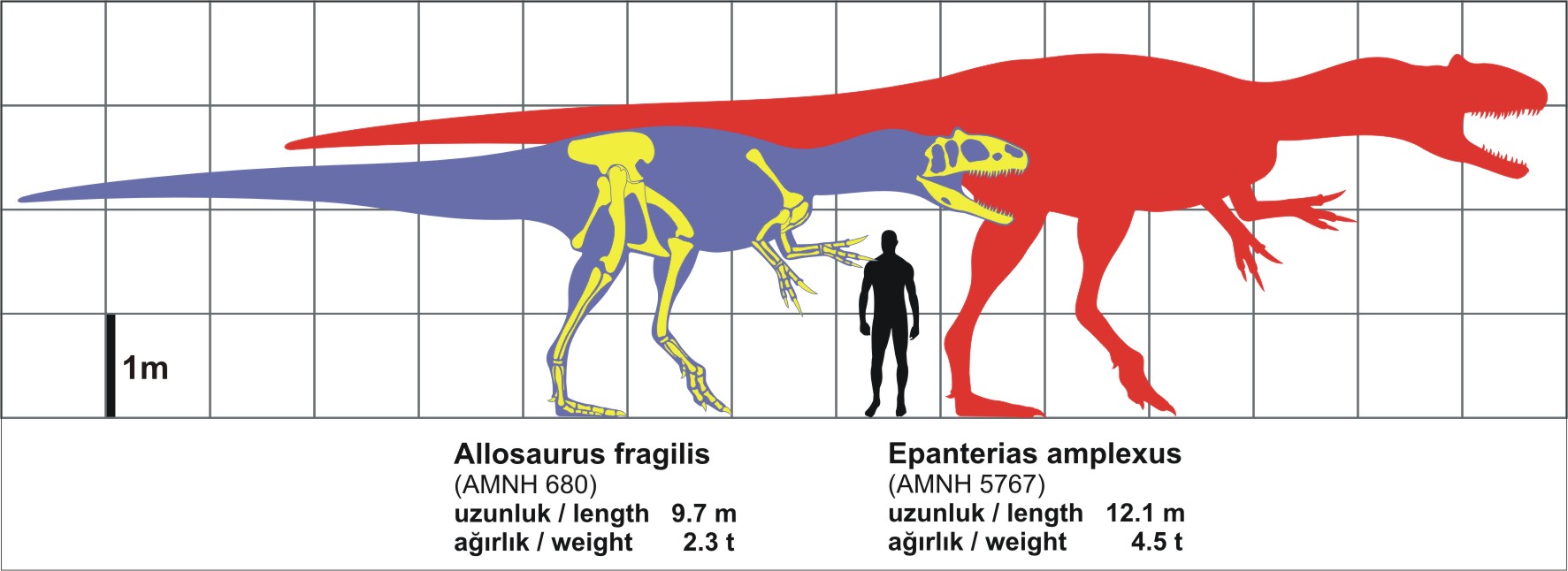
Epanterias is a dubious genus of theropod dinosaur from the Kimmeridgian-Tithonian-age Upper Jurassic upper Morrison Formation of Garden Park, Colorado. It was described by Edward Drinker Cope in 1878. The type species is Epanterias amplexus.This genus is based on what is now AMNH 5767, parts of three vertebrae, a coracoid, and a metatarsal.Although Cope thought it was a sauropod it was later shown to be a theropod.Gregory S. Paul reassessed the material as pertaining to a large species of Allosaurus in 1988 (which he classified as Allosaurus amplexus).Other authors have gone further and considered E. amplexus as simply a large individual of Allosaurus fragilis.In 2010, Gregory S. Paul and Kenneth Carpenter noted that the E. amplexus specimen comes from higher in the Morrison Formation than the type specimen of Allosaurus fragilis, and is therefore "probably a different taxon". They also considered its holotype specimen not diagnostic and classified it as a nomen dubium
Siats meekerorum

Siats is an extinct genus of large megaraptoran theropod dinosaur known from the Late Cretaceous Cedar Mountain Formation of Utah, US. It contains a single species, Siats meekerorum. S. meekerorum would be the first neovenatorid discovered in North America and the geologically youngest allosauroid yet discovered from the continent.However, recent phylogenetic analysis suggests that all megaraptorans are actually tyrannosauroids.
Siats was first described and named by Lindsay E. Zanno and Peter J. Makovicky in 2013 and the type species is Siats meekerorum. The generic name is derived from the name of Siats, a man-eating monster in the Ute mythology. The specific name meekerorum honours the late geologist John Caldwell Meeker who bequeathed a fund for the support of paleontological research, his widow Withrow Meeker and their daughter Lis Meeker, one of the volunteers in the research project.
Siats is known from the holotype FMNH PR 2716, a partial postcranial skeleton housed at the Field Museum of Natural History, Chicago. FMNH PR 2716 consists of five dorsal and eight caudal vertebrae, a chevron, partial right ilium, ischium and fibula, a partial left tibia, and several right and left pedal phalanges. FMNH PR 2716 was discovered by Lindsay Zanno, as a part of a 2008 expedition of the Field Museum led by Peter Makovicky. It was collected between 2008 and 2010 from the Mussentuchit Member of the Cedar Mountain Formation, in Emery County of Utah, dating to the early Cenomanian stage of the Late Cretaceous, approximately 98.5 million years ago.
The holotype came from a single immature individual, based on the incomplete fusion of neural arches to their centra in the vertebral column. Siats is characterized by seven diagnostic, including four autapomorphic (i.e. unique), traits. Its autapomorphies include the subtriangular cross section of the distal caudal vertebrae, elongated centrodiapophyseal laminae lacking noticeable infradiapophyseal fossae on the proximal caudals, a transversely concaved acetabular rim of iliac pubic peduncle, and the presence of a notch on the end of the truncated lateral brevis shelf. Other notable traits include the broad neural spines on the dorsal vertebrae. Siats represents one of the largest known theropods from North America. Zanno and Mackovicky (2013) using other Megaraptoran taxa as a proxy, estimate the femur length of the holotype specimen FMNH PR 2716 at 1.19 meters long, and using a femur circumference regression, estimate its mass at roughly 4 tons. Zanno and Mackovicky also wrote that the holotype specimen was already comparable in size to Saurophaganax and Acrocanthosaurus despite the visible neurocentral sutures showing absence of ossification, suggesting a skeletally immature individual.
Тагове:
Encyclopedia Largest Prehistoric Animals...
Encyclopedia Largest prehistoric animals...
