
Постинг
20.12.2015 19:12 -
Encyclopedia Largest prehistoric animals Vol.1 Vertebrates part2/3 Raptors - Hollywood nonsense
Автор: valentint
Категория: Забавление

Прочетен: 5542 Коментари: 0 Гласове:
Последна промяна: 14.06.2019 21:03
Прочетен: 5542 Коментари: 0 Гласове:
0
Последна промяна: 14.06.2019 21:03
Velociraptor
The terrible predator of Hollywood movies was not more dangerous to humans than the turkey we eat at Christmas

Earlier that year, Osborn had called the dinosaur Ovoraptor djadochtari in an article in the popular press, but the creature wasn"t formally described in the article and the name "Ovoraptor" wasn"t mentioned in a scientific journal, making Velociraptor the accepted name.
There are two Velociraptor species, V. mongoliensis and V. osmolskae, the second of which was only identified in 2008.
A member of the Dromaeosauridae family of small- to medium-sized birdlike dinosaurs, Velociraptor was roughly the size of a small turkey and smaller than others in this family of dinosaurs, which included Deinonychus and Achillobator. Adult Velociraptors grew up to 6.8 feet (2 meters) long, 1.6 feet (0.5 meter) tall at the hip and weighed up to 33 lbs. (15 kilograms).
Like Tyrannosaurus rex, Velociraptor had a prominent role in the "Jurassic Park" movies, but scientists do not believe it resembled anything close to its Hollywood depiction in terms of size or appearance. In fact, the movies" Velociraptor was actually modeled after Deinonychus, and sported a similar size and snout.
While the Velociraptor was featherless in the movies, paleontologists discovered quill knobs (places where the flight-related feathers of birds are anchored to the bone) on a well-preserved Velociraptor forearm from Mongolia in 2007, indicating the dinosaur had feathers.
Despite having feathers, however, the arms of Velociraptors were too short to allow them to fly or even glide. The find suggests that the dinosaurs" dromaeosaurid ancestors could fly at one point, but lost that ability, according to the study published in the journal Science.
Velociraptor retained its feathers, and possibly used them to attract mates, regulate body temperature, protect eggs from the environment or generate thrust and speed while running up inclines.
Velociraptor had a relatively large skull , which was about 9.1 inches (23 centimeters) long, concave on the upper surface and convex on the lower surface, according to a 1999 description of a Velociraptor skull, published in the journal Acta Palaeontologica Polonica. Additionally, its snout was long, narrow and shallow, and made up about 60 percent of the dinosaur"s entire skull length.
Velociraptor had 13 to 15 teeth in its upper jaw and 14 to 15 teeth in its lower jaw. These teeth were widely spaced and serrated, though more strongly on the back edge than the front.
Velociraptor"s tail of hard, fused bones was inflexible, but likely kept it balanced as it ran, hunted and jumped.
Velociraptor, like other dromaeosaurids, had two large hand-like appendages with three curved claws. They also had a sickle-shaped talon on the second toe of each foot. They normally kept these talons off the ground like folded switchblades, and used them as hooks to keep their prey from escaping (similar to modern birds of prey), according to a study published in 2011 in the journal PLOS ONE.
What did Velociraptor eat?
Velociraptor was a carnivore that hunted and scavenged for food. "It spent the vast majority of the time eating small things," which likely included reptiles, amphibians, insects, small dinosaurs and mammals, said David Hone, a paleontologist at Queen Mary University of London.
Velociraptor, which means “speedy thief,” had a sharp, deadly, sickle-shaped, retractable, 3.5-inch (9 cm) claw on each foot (located on each second toe). The Velociraptor may have been able to run up to roughly 40 mph (60 km/hr) for short bursts. This predator may have hunted in packs.The fast predator also appears to have had a complicated relationship with Protoceratops, a sheep-sized herbivore and ancestor to Triceratops. In 1971, a Polish-Mongolian team discovered the famous "Fighting Dinosaurs" specimen — fossils of a Velociraptor and Protoceratops locked in a death grip, in which the Velociraptor embedded one of its foot claws into the neck of the Protoceratops while the Protoceratops bit down on (and probably broke) one of the Velociraptor"s arms.
Preserved in sand deposits after being buried from a collapsing sand dune or sudden sandstorm, the pair proved that Velociraptors hunted for food, but an attack on such a large animal probably wasn"t common. "Few predators ever take on prey bigger than 50 percent of their body mass," Hone told Live Science, adding that the Velociraptor could have been starving or simply "young and dumb."
But that"s not to say Velociraptor didn"t frequently eat Protoceratops carcasses. In 2008, researchers unearthed Protoceratops fossils marred with marks and grooves matching raptor teeth, as well as two teeth that belonged either to Velociraptor or another dromaeosaurid.
After analyzing the remains, Hone and his colleagues determined that the raptor didn"t kill the plant-eater. Instead, it fed on the Protoceratops, which likely had little meat left on it (hence the bite marks on the herbivore"s jaws and raptor"s knocked-out teeth), according to the study, published 2010 in the journal Palaeogeography, Palaeoclimatology, Palaeoecology.
The first Velociraptor fossil was discovered by Peter Kaisen on the first American Museum of Natural History expedition to the Outer Mongolian Gobi Desert in August 1923. The fossil consisted of a skull that was crushed but complete and a toe claw.
Velociraptor fossils have been found in the Gobi Desert, which covers southern Mongolia and parts of northern China. Velociraptor mongoliensis have only been discovered in the Djadochta (Djadokhta) Formation, which is in the Mongolian province of Цmnцgovi.
Velociraptor osmolskae was discovered at the Bayan Mandahu Formation in Inner Mongolia, China. The species was described based on a partial adult skull. [Image Gallery: Dinosaur Fossils]
Like the "Fighting Dinosaurs," other Velociraptor fossils were found in arid sand dune environments.
Troodontids
Smart dinosaurs?! - fantasies
.jpg)
Remarkably when Troodon was first "discovered" it was only known from a tooth, and it was the description of this tooth that gave rise to the genus. Although naming a creature after only discovering its teeth is hard to imagine today, it was a common place practice in the nineteenth century, and at least in the case of Troodon the teeth were different to most others.
Two problems arose from describing Troodon from only teeth. One, while these teeth were curved like in a carnivore, they had curved serrations similar to some herbivores. Two, because no one knew of any skeletal remains, no one knew what kind of dinosaur it actually was, or even if it really was a dinosaur for that matter.
When named by Joseph Leidy in 1856, Troodon was classed as a lizard. When classed as a dinosaur in 1901, Troodon was assigned to the megalosauridae, a wastebasket group at the time that was used as a dumping ground for any carnivorous dinosaur whose kind and position among others was not known. In 1924 Troodon "became" a pachycephalosaur, a herbivorous kind of dinosaur known for having dome headed skulls, with the grouping made on the basis of similarities between their teeth, notably the serrations. It should also be mentioned that as Troodon was grouped with them, all pachycephalosaurs known at the time were classed under troodontidae, until 1945 when Troodon was removed from the group on the basis that its teeth were for a carnivorous purpose. Because the group could not be called after a dinosaur that was no longer a part of it, they were re-named the pachycephalosuars after Pachycephalosaurus.
It turns out that while some of above was unfolding, skeletal material for Troodon did actually exist, although it was classified under the name of Stenonychosaurus. The material for Stenonychosaurus was discovered in 1932 but was fragmentary, and when compared to the zero skeletal remains of Troodon, no one knew any better than to create a separate genus. The discovery of more fossil material throughout the twentieth century would slowly bring Stenonychosaurus and Troodon together, but it would not be until 1987 when Dr Philip Currie studied material attributed to both that the connection was made. Currie found that the only differences between Stenonychosaurus and Troodon were the products of the age of the individuals and they were one and the same. Not only did Stenonychosaurus become a junior synonym, but Polydontosaurus and Pectinodon also became junior synonyms of Troodon.
Not long after this reclassification occurred many, including Currie himself, questioned if it was actually correct to name just one species of Troodon. This is based upon the facts that Troodon specimens are separated by large geographic distances as well as several million years in the fossil record. With this in mind there have been calls to re-examine all Troodon fossils, to see how many species there actually are. The problem here returns to the fact that the holotype specimen that all subsequent fossil finds are compared to is a only a tooth. The only possible way to sort things out with Troodon remaining a valid name and not a Nomen dubium would be establish a neotype specimen. This is where more complete material is set as the "standard" for a species so that future material can be compared against it for the purposes of identification. The establishment of a neotype is not unknown and had to be done with another well-known dinosaur, Coelophysis.

Although small for a dinosaur, Troodon is still the largest member of the troodontidae group. Troodon also had large eyes that faced forwards allowing Troodon to make use of stereoscopic vision. Aside from having a good degree of depth perception, the eyes may well have also allowed for exceptional night vision due to their large size.
The characteristic teeth of Troodon do not make its diet easy to establish. Although overall they are recurved like those of a predator, they have serrations that suggest herbivore. What can be gathered is this; as a Theropod, Troodon had carnivorous ancestors, and this is confirmed in the recurved shape of the teeth. Recurved teeth curve towards the tip so that they point to the back of the mouth. This design is a common trait in predators and exists so that seized prey cannot escape.
The teeth serrations however are more like those of a herbivore that specialises in soft vegetation, and similar serrations can be seen in the teeth of herbivorous dinosaurs such as Pachycephalosaurus and even Stegosaurus. Either the serrations are an "evolutionary experiment", or Troodon possibly included some plant material into its diet. One thing that is certain about the teeth, they would have been almost useless against hard material like bone, or even tough branches and Troodon would have had to focus on only softer food items.
The snout on Troodon has a "U" shape to it when viewed from above. This has been suggested as an herbivorous trait as it is often seen in herbivorous lizards. It should be remembered though that one of the most famous carnivorous dinosaurs of all time, Tyrannosaurus, also had a "U" shaped snout, an adaptation that allowed it to take bigger bites.
When considering the other predatory traits such as forward facing eyes, long legs, and light weight build and the presence of a sickle claw on each foot, it is almost certain that Troodon was a predator. Even with this reasoning however, it is still possible that Troodon may have been an occasional omnivore, should Troodon have found the right food item.
Another feature that makes Troodon stand out from other dinosaurs is the presence of an opposable finger. Although not constructed like a human hand, the best way to describe it is if you try grasping something between your thumb and just two of your fingers. It is unknown how much use Troodon would have given this adaptation, but the fact that it developed in the first place would suggest that there was at least one good reason for it occurring. It may have been to better manipulate ground objects or moving undergrowth branches when searching for prey.
While old depictions of Troodon (then known as Stenonychosaurus) show a lizard like dinosaur with bare skin, most modern reconstructions place a covering of primitive feathers over its body. This is quite a reasonable supposition as not only did other smaller members of the troodontidae have feathers, Troodon is known from as far north as modern day Alaska. Such a northern latitude even at the end of the Cretaceous period would have been much cooler than areas just a few hundred miles south, quite easily necessitating the presence of insulating feathers.
The discovery of fossilised nest sites in the Two Medicine Formation has revealed very interesting insights into how Troodon laid and brooded over their eggs. Troodon nests were dish shaped with a one meter diameter. The nests seem to have been built from just sediment with no other material in the construction. The nests examined show evidence of between sixteen and twenty-four eggs per nest with some seen to have hatched.
The eggs themselves are like elongated teardrops. When laid, the pointed end is stuck into the sediment, and is presumably shaped for this purpose. The eggs are also laid at an angle pointing towards the centre of the nest. Because eggs are grouped into pairs, it is thought that in life, Troodon would have had two oviducts, something which is seen in modern crocodiles.
Different nest sites reveal different stages in the incubation, but all point to the same conclusion. Nests are not found where only a few of the eggs have hatched, either they all are, or none at all. Also analysis of fossilised embryos in unhatched eggs of the same nest show that they are all at the same developmental stage. It would probably be impossible for a Troodon to lay up to two dozen eggs in one go, so it is thought that the eggs were laid two at a time. The main brooding did not begin until all of the eggs had been laid so that all of the young could develop and more importantly hatch at the same time as one another.
One nest site was also found with the remains of an adult Troodon. Analysis showed that this individual was a male, based upon the fact that is did not have the same bone resorption patterns as would be expected in a female during ovulation. This has raised the notion that male Troodon had an active role in the brooding of the nest as can be seen in many of today’s birds.
Because of its large brain size relative to its overall body mass. Troodon is considered to quite possibly be the smartest dinosaur known to exist. This is based upon the brain to body mass ratio, and sometimes the encephalization quotient. The encephalization quotient is considered the more accurate of the two as it takes the relative size of the animal into account. Both systems work by comparing the size of the brain to the size of the animal.
Although both of the systems seem quite complex to understand, they actually run on a pretty basic principal. Neural cells that make up the brain are the same size regardless of the size of the creature. This means that a larger brain is made up of more cells, and more cells is taken to mean more brain power. An animal with a larger body may need more of this power for motor skills (moving of muscles), and with this taken into account, the general intelligence of the animal is estimated.
The problem with both systems is that neither takes into account the type of neural tissue. Brains are made up of different areas, sight, smell, memory, etc. If two different animals still had the same size brain as one another, one still may be more intelligent than the other if one had more neural tissue associated with things like memory and problem solving than sight and smell. Although that animal would not be as physically able as the other, it would have greater intelligence.
Back to the subject of intelligence in Troodon, interpretation and re-creation of the brain have been key areas of study. On the EQ scale, mankind places about 7.4-7.8. Some studies have placed Troodon as high as 5.8, quite an achievement when you consider that most dinosaurs chart under 2 on the scale, with most herbivorous dinosaurs under 1. Modern animals like a Chimpanzee chart around 2.2-2.5, and Bottlenose Dolphins just over 4.
As stated above it is the kind of tissue that matters. Troodon had large eyes, and coupled with its presence in Northern latitudes, may have needed more brain power just to process visual information. However this does not disprove Troodon intelligence. Brains in higher forms, including people are "folded". This is the wrinkled look of brains when they are seen outside of the body, and occurs in order to get more neural tissue working in a smaller area. The brains of more primitive animals are usually just amorphous clumps of neural cells. Reconstruction of Troodon brains has revealed early indications of the brain beginning to fold on itself, something that is considered to be radically different to other reconstructed dinosaur brains.
Again this does not mean that Troodon was more intelligent, but it does indicate that it had more potential brain power that was developing along even more special lines. It is also worth remembering that by having hands that could grasp things, Troodon would have had even greater opportunity to learn from its environment.
The idea of dinosaurs with increasing intelligence and dexterous hands has led many too wonder how dinosaurs like Troodon would have evolved had the KT extinction not occurred sixty-five million years ago. Well in 1982 a man named Dale Russell put form to the thought. Based upon Troodon ( then still known as Stenonychosaurus), Russell presented a humanoid dinosaur, which means that it was depicted as walking in a similar stance and posture as a human being. The body proportions were also close to that of a human. The eyes were still large, and the Dinosauroid was envisioned as giving birth to live young and feeding them upon regurgitated food.
Although some have criticised the reconstruction on the grounds that it is "too human", and descendants would have probably retained a more "classic" theropod body, it is somewhat missing the point as the Dinosauroid reconstruction is more of a "what if" than an absolute. It is still worth thinking about though that mankind is thought to have evolved from apes over the course of hundreds of thousands of years. Imagine what a dinosaur already smarter than a Chimpanzee could do over sixty-five million.
The terrible predator of Hollywood movies was not more dangerous to humans than the turkey we eat at Christmas
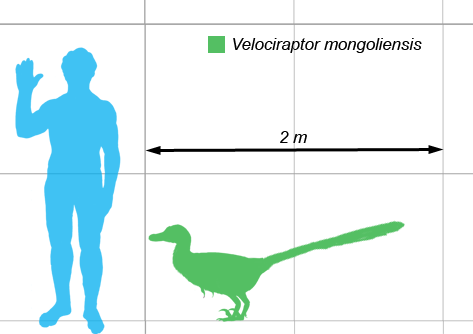
Although it featured some nasty looking claws, the Velociraptor was actually a lot smaller than it was portrayed in the movie Jurassic Park, more the size of a turkey than a human. Velociraptor was a mid-sized dromaeosaurid, with adults measuring up to 2.07 m (6.8 ft) long, 0.5 m (1.6 ft) high at the hip, and weighing up to 15 kg (33 lb).The skull, which grew up to 25 cm (10 in) long, was uniquely up-curved, concave on the upper surface and convex on the lower. The jaws were lined with 26–28 widely spaced teeth on each side, each more strongly serrated on the back edge than the front.
Velociraptor roamed the Earth about 85.8 million to 70.6 million years ago during the end of the Cretaceous Periiod.
Earlier that year, Osborn had called the dinosaur Ovoraptor djadochtari in an article in the popular press, but the creature wasn"t formally described in the article and the name "Ovoraptor" wasn"t mentioned in a scientific journal, making Velociraptor the accepted name.
There are two Velociraptor species, V. mongoliensis and V. osmolskae, the second of which was only identified in 2008.
A member of the Dromaeosauridae family of small- to medium-sized birdlike dinosaurs, Velociraptor was roughly the size of a small turkey and smaller than others in this family of dinosaurs, which included Deinonychus and Achillobator. Adult Velociraptors grew up to 6.8 feet (2 meters) long, 1.6 feet (0.5 meter) tall at the hip and weighed up to 33 lbs. (15 kilograms).
Like Tyrannosaurus rex, Velociraptor had a prominent role in the "Jurassic Park" movies, but scientists do not believe it resembled anything close to its Hollywood depiction in terms of size or appearance. In fact, the movies" Velociraptor was actually modeled after Deinonychus, and sported a similar size and snout.
While the Velociraptor was featherless in the movies, paleontologists discovered quill knobs (places where the flight-related feathers of birds are anchored to the bone) on a well-preserved Velociraptor forearm from Mongolia in 2007, indicating the dinosaur had feathers.
Despite having feathers, however, the arms of Velociraptors were too short to allow them to fly or even glide. The find suggests that the dinosaurs" dromaeosaurid ancestors could fly at one point, but lost that ability, according to the study published in the journal Science.
Velociraptor retained its feathers, and possibly used them to attract mates, regulate body temperature, protect eggs from the environment or generate thrust and speed while running up inclines.
Velociraptor had a relatively large skull , which was about 9.1 inches (23 centimeters) long, concave on the upper surface and convex on the lower surface, according to a 1999 description of a Velociraptor skull, published in the journal Acta Palaeontologica Polonica. Additionally, its snout was long, narrow and shallow, and made up about 60 percent of the dinosaur"s entire skull length.
Velociraptor had 13 to 15 teeth in its upper jaw and 14 to 15 teeth in its lower jaw. These teeth were widely spaced and serrated, though more strongly on the back edge than the front.
Velociraptor"s tail of hard, fused bones was inflexible, but likely kept it balanced as it ran, hunted and jumped.
Velociraptor, like other dromaeosaurids, had two large hand-like appendages with three curved claws. They also had a sickle-shaped talon on the second toe of each foot. They normally kept these talons off the ground like folded switchblades, and used them as hooks to keep their prey from escaping (similar to modern birds of prey), according to a study published in 2011 in the journal PLOS ONE.
What did Velociraptor eat?
Velociraptor was a carnivore that hunted and scavenged for food. "It spent the vast majority of the time eating small things," which likely included reptiles, amphibians, insects, small dinosaurs and mammals, said David Hone, a paleontologist at Queen Mary University of London.
Velociraptor, which means “speedy thief,” had a sharp, deadly, sickle-shaped, retractable, 3.5-inch (9 cm) claw on each foot (located on each second toe). The Velociraptor may have been able to run up to roughly 40 mph (60 km/hr) for short bursts. This predator may have hunted in packs.The fast predator also appears to have had a complicated relationship with Protoceratops, a sheep-sized herbivore and ancestor to Triceratops. In 1971, a Polish-Mongolian team discovered the famous "Fighting Dinosaurs" specimen — fossils of a Velociraptor and Protoceratops locked in a death grip, in which the Velociraptor embedded one of its foot claws into the neck of the Protoceratops while the Protoceratops bit down on (and probably broke) one of the Velociraptor"s arms.
Preserved in sand deposits after being buried from a collapsing sand dune or sudden sandstorm, the pair proved that Velociraptors hunted for food, but an attack on such a large animal probably wasn"t common. "Few predators ever take on prey bigger than 50 percent of their body mass," Hone told Live Science, adding that the Velociraptor could have been starving or simply "young and dumb."
But that"s not to say Velociraptor didn"t frequently eat Protoceratops carcasses. In 2008, researchers unearthed Protoceratops fossils marred with marks and grooves matching raptor teeth, as well as two teeth that belonged either to Velociraptor or another dromaeosaurid.
After analyzing the remains, Hone and his colleagues determined that the raptor didn"t kill the plant-eater. Instead, it fed on the Protoceratops, which likely had little meat left on it (hence the bite marks on the herbivore"s jaws and raptor"s knocked-out teeth), according to the study, published 2010 in the journal Palaeogeography, Palaeoclimatology, Palaeoecology.
The first Velociraptor fossil was discovered by Peter Kaisen on the first American Museum of Natural History expedition to the Outer Mongolian Gobi Desert in August 1923. The fossil consisted of a skull that was crushed but complete and a toe claw.
Velociraptor fossils have been found in the Gobi Desert, which covers southern Mongolia and parts of northern China. Velociraptor mongoliensis have only been discovered in the Djadochta (Djadokhta) Formation, which is in the Mongolian province of Цmnцgovi.
Velociraptor osmolskae was discovered at the Bayan Mandahu Formation in Inner Mongolia, China. The species was described based on a partial adult skull. [Image Gallery: Dinosaur Fossils]
Like the "Fighting Dinosaurs," other Velociraptor fossils were found in arid sand dune environments.
Troodontids
Smart dinosaurs?! - fantasies
.jpg)
Remarkably when Troodon was first "discovered" it was only known from a tooth, and it was the description of this tooth that gave rise to the genus. Although naming a creature after only discovering its teeth is hard to imagine today, it was a common place practice in the nineteenth century, and at least in the case of Troodon the teeth were different to most others.
Two problems arose from describing Troodon from only teeth. One, while these teeth were curved like in a carnivore, they had curved serrations similar to some herbivores. Two, because no one knew of any skeletal remains, no one knew what kind of dinosaur it actually was, or even if it really was a dinosaur for that matter.
When named by Joseph Leidy in 1856, Troodon was classed as a lizard. When classed as a dinosaur in 1901, Troodon was assigned to the megalosauridae, a wastebasket group at the time that was used as a dumping ground for any carnivorous dinosaur whose kind and position among others was not known. In 1924 Troodon "became" a pachycephalosaur, a herbivorous kind of dinosaur known for having dome headed skulls, with the grouping made on the basis of similarities between their teeth, notably the serrations. It should also be mentioned that as Troodon was grouped with them, all pachycephalosaurs known at the time were classed under troodontidae, until 1945 when Troodon was removed from the group on the basis that its teeth were for a carnivorous purpose. Because the group could not be called after a dinosaur that was no longer a part of it, they were re-named the pachycephalosuars after Pachycephalosaurus.
It turns out that while some of above was unfolding, skeletal material for Troodon did actually exist, although it was classified under the name of Stenonychosaurus. The material for Stenonychosaurus was discovered in 1932 but was fragmentary, and when compared to the zero skeletal remains of Troodon, no one knew any better than to create a separate genus. The discovery of more fossil material throughout the twentieth century would slowly bring Stenonychosaurus and Troodon together, but it would not be until 1987 when Dr Philip Currie studied material attributed to both that the connection was made. Currie found that the only differences between Stenonychosaurus and Troodon were the products of the age of the individuals and they were one and the same. Not only did Stenonychosaurus become a junior synonym, but Polydontosaurus and Pectinodon also became junior synonyms of Troodon.
Not long after this reclassification occurred many, including Currie himself, questioned if it was actually correct to name just one species of Troodon. This is based upon the facts that Troodon specimens are separated by large geographic distances as well as several million years in the fossil record. With this in mind there have been calls to re-examine all Troodon fossils, to see how many species there actually are. The problem here returns to the fact that the holotype specimen that all subsequent fossil finds are compared to is a only a tooth. The only possible way to sort things out with Troodon remaining a valid name and not a Nomen dubium would be establish a neotype specimen. This is where more complete material is set as the "standard" for a species so that future material can be compared against it for the purposes of identification. The establishment of a neotype is not unknown and had to be done with another well-known dinosaur, Coelophysis.
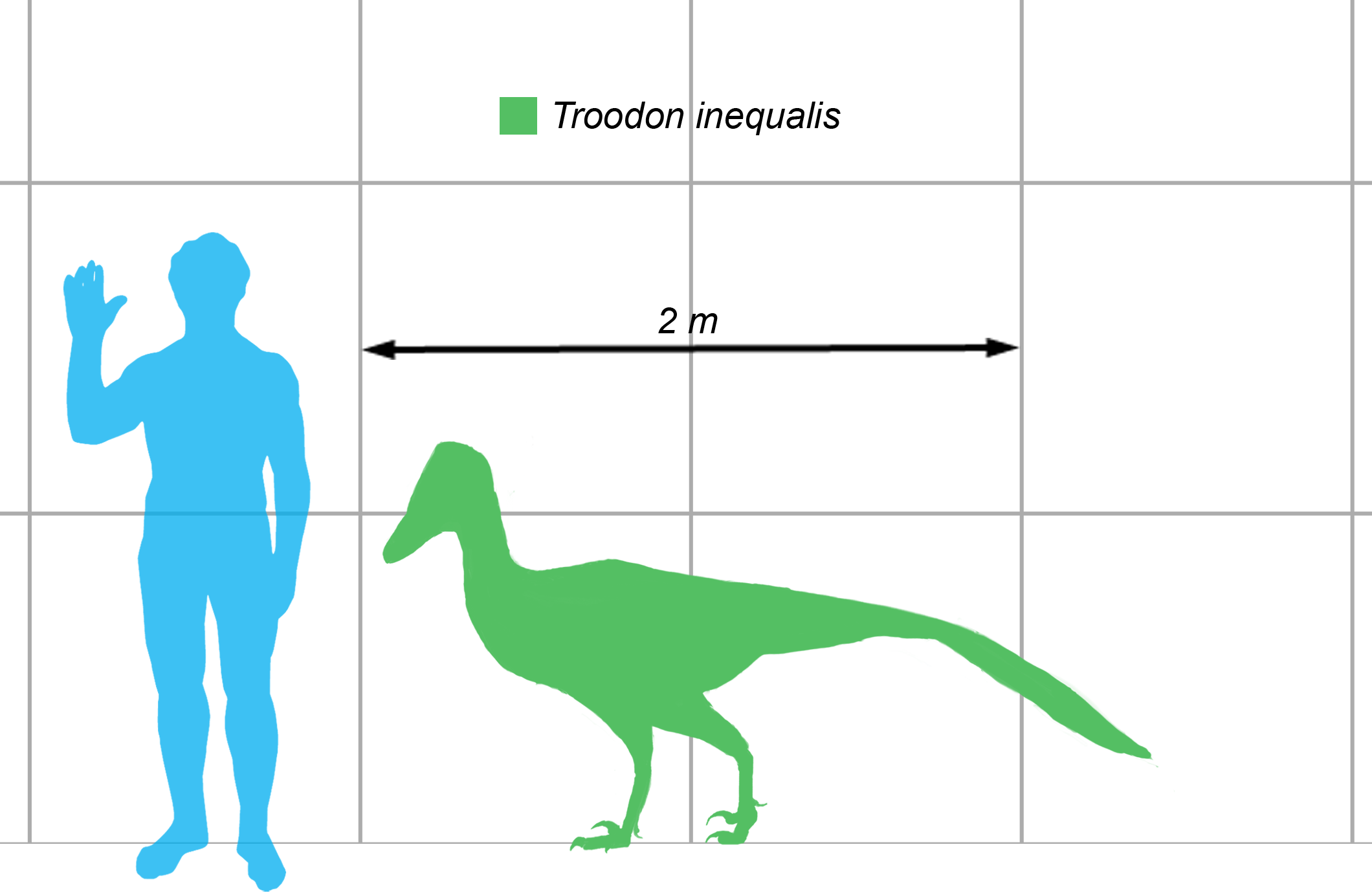
Although small for a dinosaur, Troodon is still the largest member of the troodontidae group. Troodon also had large eyes that faced forwards allowing Troodon to make use of stereoscopic vision. Aside from having a good degree of depth perception, the eyes may well have also allowed for exceptional night vision due to their large size.
The characteristic teeth of Troodon do not make its diet easy to establish. Although overall they are recurved like those of a predator, they have serrations that suggest herbivore. What can be gathered is this; as a Theropod, Troodon had carnivorous ancestors, and this is confirmed in the recurved shape of the teeth. Recurved teeth curve towards the tip so that they point to the back of the mouth. This design is a common trait in predators and exists so that seized prey cannot escape.
The teeth serrations however are more like those of a herbivore that specialises in soft vegetation, and similar serrations can be seen in the teeth of herbivorous dinosaurs such as Pachycephalosaurus and even Stegosaurus. Either the serrations are an "evolutionary experiment", or Troodon possibly included some plant material into its diet. One thing that is certain about the teeth, they would have been almost useless against hard material like bone, or even tough branches and Troodon would have had to focus on only softer food items.
The snout on Troodon has a "U" shape to it when viewed from above. This has been suggested as an herbivorous trait as it is often seen in herbivorous lizards. It should be remembered though that one of the most famous carnivorous dinosaurs of all time, Tyrannosaurus, also had a "U" shaped snout, an adaptation that allowed it to take bigger bites.
When considering the other predatory traits such as forward facing eyes, long legs, and light weight build and the presence of a sickle claw on each foot, it is almost certain that Troodon was a predator. Even with this reasoning however, it is still possible that Troodon may have been an occasional omnivore, should Troodon have found the right food item.
Another feature that makes Troodon stand out from other dinosaurs is the presence of an opposable finger. Although not constructed like a human hand, the best way to describe it is if you try grasping something between your thumb and just two of your fingers. It is unknown how much use Troodon would have given this adaptation, but the fact that it developed in the first place would suggest that there was at least one good reason for it occurring. It may have been to better manipulate ground objects or moving undergrowth branches when searching for prey.
While old depictions of Troodon (then known as Stenonychosaurus) show a lizard like dinosaur with bare skin, most modern reconstructions place a covering of primitive feathers over its body. This is quite a reasonable supposition as not only did other smaller members of the troodontidae have feathers, Troodon is known from as far north as modern day Alaska. Such a northern latitude even at the end of the Cretaceous period would have been much cooler than areas just a few hundred miles south, quite easily necessitating the presence of insulating feathers.
The discovery of fossilised nest sites in the Two Medicine Formation has revealed very interesting insights into how Troodon laid and brooded over their eggs. Troodon nests were dish shaped with a one meter diameter. The nests seem to have been built from just sediment with no other material in the construction. The nests examined show evidence of between sixteen and twenty-four eggs per nest with some seen to have hatched.
The eggs themselves are like elongated teardrops. When laid, the pointed end is stuck into the sediment, and is presumably shaped for this purpose. The eggs are also laid at an angle pointing towards the centre of the nest. Because eggs are grouped into pairs, it is thought that in life, Troodon would have had two oviducts, something which is seen in modern crocodiles.
Different nest sites reveal different stages in the incubation, but all point to the same conclusion. Nests are not found where only a few of the eggs have hatched, either they all are, or none at all. Also analysis of fossilised embryos in unhatched eggs of the same nest show that they are all at the same developmental stage. It would probably be impossible for a Troodon to lay up to two dozen eggs in one go, so it is thought that the eggs were laid two at a time. The main brooding did not begin until all of the eggs had been laid so that all of the young could develop and more importantly hatch at the same time as one another.
One nest site was also found with the remains of an adult Troodon. Analysis showed that this individual was a male, based upon the fact that is did not have the same bone resorption patterns as would be expected in a female during ovulation. This has raised the notion that male Troodon had an active role in the brooding of the nest as can be seen in many of today’s birds.
Because of its large brain size relative to its overall body mass. Troodon is considered to quite possibly be the smartest dinosaur known to exist. This is based upon the brain to body mass ratio, and sometimes the encephalization quotient. The encephalization quotient is considered the more accurate of the two as it takes the relative size of the animal into account. Both systems work by comparing the size of the brain to the size of the animal.
Although both of the systems seem quite complex to understand, they actually run on a pretty basic principal. Neural cells that make up the brain are the same size regardless of the size of the creature. This means that a larger brain is made up of more cells, and more cells is taken to mean more brain power. An animal with a larger body may need more of this power for motor skills (moving of muscles), and with this taken into account, the general intelligence of the animal is estimated.
The problem with both systems is that neither takes into account the type of neural tissue. Brains are made up of different areas, sight, smell, memory, etc. If two different animals still had the same size brain as one another, one still may be more intelligent than the other if one had more neural tissue associated with things like memory and problem solving than sight and smell. Although that animal would not be as physically able as the other, it would have greater intelligence.
Back to the subject of intelligence in Troodon, interpretation and re-creation of the brain have been key areas of study. On the EQ scale, mankind places about 7.4-7.8. Some studies have placed Troodon as high as 5.8, quite an achievement when you consider that most dinosaurs chart under 2 on the scale, with most herbivorous dinosaurs under 1. Modern animals like a Chimpanzee chart around 2.2-2.5, and Bottlenose Dolphins just over 4.
As stated above it is the kind of tissue that matters. Troodon had large eyes, and coupled with its presence in Northern latitudes, may have needed more brain power just to process visual information. However this does not disprove Troodon intelligence. Brains in higher forms, including people are "folded". This is the wrinkled look of brains when they are seen outside of the body, and occurs in order to get more neural tissue working in a smaller area. The brains of more primitive animals are usually just amorphous clumps of neural cells. Reconstruction of Troodon brains has revealed early indications of the brain beginning to fold on itself, something that is considered to be radically different to other reconstructed dinosaur brains.
Again this does not mean that Troodon was more intelligent, but it does indicate that it had more potential brain power that was developing along even more special lines. It is also worth remembering that by having hands that could grasp things, Troodon would have had even greater opportunity to learn from its environment.
The idea of dinosaurs with increasing intelligence and dexterous hands has led many too wonder how dinosaurs like Troodon would have evolved had the KT extinction not occurred sixty-five million years ago. Well in 1982 a man named Dale Russell put form to the thought. Based upon Troodon ( then still known as Stenonychosaurus), Russell presented a humanoid dinosaur, which means that it was depicted as walking in a similar stance and posture as a human being. The body proportions were also close to that of a human. The eyes were still large, and the Dinosauroid was envisioned as giving birth to live young and feeding them upon regurgitated food.
Although some have criticised the reconstruction on the grounds that it is "too human", and descendants would have probably retained a more "classic" theropod body, it is somewhat missing the point as the Dinosauroid reconstruction is more of a "what if" than an absolute. It is still worth thinking about though that mankind is thought to have evolved from apes over the course of hundreds of thousands of years. Imagine what a dinosaur already smarter than a Chimpanzee could do over sixty-five million.
Тагове:
Encyclopedia Largest prehistoric animals...
Encyclopedia Largest prehistoric animals...
Encyclopedia Largest Prehistoric Animals...
Encyclopedia Largest prehistoric animals...
Encyclopedia Largest Prehistoric Animals...
Следващ постинг
Предишен постинг
Няма коментари
