

Прочетен: 6854 Коментари: 0 Гласове:
Последна промяна: 05.07.2019 21:54

1. Tachyglossus 2. Megalibgwilia 3. Zaglossus
Monotremata is the most ancient living order of mammals. In addition to being egg layers (oviparous), members of this order share primitive skeletal features such as the shoulder girdle and skull characteristics that have been lost in other living mammals. Relationships to other mammalian groups have been difficult to determine because of the puzzling combination of primitive features and specialized characteristics, a phenomenon known as mosaic evolution. Certain features of the skull appear to link monotremes to the extinct early mammal groups. Other evidence, particularly genetic data, places Monotremata close to more-advanced mammals, namely marsupials.
Monotremes have some features that may be inherited from the cynodont ancestors:
1.like lizards and birds, they use the same orifice to urinate, defecate and reproduce ("monotreme" means "one hole").
2.they lay eggs that are leathery and uncalcified, like those of lizards, turtles and crocodilians.
Unlike other mammals, female monotremes do not have nipples and feed their young by "sweating" milk from patches on their bellies.
These features are not visible in fossils, and the main characteristics from paleontologists" point of view are:
1.A slender dentary bone in which the coronoid process is small or non-existent.
2.The external opening of the ear lies at the posterior base of the jaw.
3.The jugal bone is small or non-existent.
4.A primitive pectoral girdle with strong ventral elements: coracoids, clavicles and interclavicle. Note: therian mammals have no interclavicle.
5.Sprawling or semi-sprawling forelimbs.
Monotremes are very highly modified for their particular ecological niches. The platypus is a semiaquatic carnivore-insectivore, and the echidnas have a specialized diet of small invertebrates such as ants, termites, and worms. The name Monotremata means “one-holed,” in reference to the fact that both sexes have only one opening at the rear of the body, which is used for both reproduction and excretion.

Various mammals of the Mesozoic Era (251 to 65.5 million years ago) with more-advanced shoulder girdles (including a multituberculate) have recently been discovered, indicating that the monotreme shoulder girdle developed before the shoulder girdles of some other mammalian groups. None of the theories of monotreme relationships has been universally accepted, although the subject is of great interest to scientists who study mammalian evolution.
Included within the monotreme order are two families known only from early in the Cretaceous Period (145.5 to 65.5 million years ago) in Australia: the platypus-like Steropodontidae, represented by a single species (Steropodon galmani), and the uniquely specialized Kollikodontidae, which is also represented by a single species (Kollikodon ritchiei). Both are known only from opalized jaw fragments. The strange rounded cusps on the molar teeth of K. ritchiei were a surprise to paleontologists, suggesting that Cretaceous monotremes may have been more diverse and widespread than previously thought and may in fact have been the dominant mammals in the Australian sector of the ancient supercontinent of Gondwanaland.The Cretaceous discoveries bring the number of families within the order to four.

1. Obdurodon 2. Steropodon 3. Teinolophos
Teinolophos, from Australia, is the earliest known monotreme. A 2007 study (published 2008) suggests that it was not a basal (primitive, ancestral) monotreme but a full-fledged platypus, and therefore that the platypus and echidna lineages diverged considerably earlier.A more recent study (2009), however, has suggested that, while Teinolophos was a type of platypus, it was also a basal monotreme and predated the radiation of modern monotremes. The semi-aquatic lifestyle of platypuses prevented them from being outcompeted by the marsupials that migrated to Australia millions of years ago, since joeys need to remain attached to their mothers and would drown if their mothers ventured into water (though there are exceptions like the water opossum and the lutrine opossum; however, they both live in South America and thus don"t come into contact with monotremes). Genetic evidence has determined that echidnas diverged from the platypus lineage as recently as 19-48M, when they made their transition from semi-aquatic to terrestrial lifestyle.
The oldest fossil platypus is recorded from the Paleogene Period (65.5 to 23 million years ago) of South America, providing evidence that during this time platypuses were distributed from Australia through Antarctica to South America.
Although the fossil record extends back to the early Cretaceous, origins of the group undoubtedly lie much farther back in time. Anatomic evidence such as the shoulder girdle suggests an origin perhaps as long ago as the middle of the Jurassic Period (199.6 to 145.5 million years ago).
Many researchers agree that the primitive features in monotremes reflect an ancient origin. Any attempt to determine the relationships of monotremes must take into account these primitive retentions. This is problematic in that either these archaic features reflect an origin far removed from that of living placental and marsupial mammals or else a mechanism must be found to explain how these features could be possessed by a group that is closely related to mammals without these traits.
The largest monotreme (egg-laying mammal) ever was the extinct long-beaked echidna species known as Zsaglosus hacketti, known from a couple of bones found in Western Australia. It was the size of a sheep, weighing probably up to 30 kg (66 lb).

Though only known from partial post cranial remains, these fossils have been reconstructed to form an echidna that in life was about one meter long. This makes Zaglossus hacketti the largest monotreme mammal to exist that we know about. Zaglossus hacketti is one species of the Zaglossus genus of echidnas, which includes species living today called the long beaked echidnas because of the shape of the snout. However, at the time of writing there are no known fossils of the skull of Zaglossus hacketti, meaning a classification within the Zaglossus genus is uncertain until a potential future fossil discovery can prove the issue one way or another.
Fossils of Zaglossus hacketti have been found with chips and burn marks upon them, indicating that the holotype individual was killed and then cooked by early aboriginal people. A combination of hunting and habitat change brought about by the arrival of the first aboriginal people in Australia has been blamed for much of the disappearance of the megafauna of Australia towards the end of the Pleistocene period. Apart from this evidence of cooking, rock art has also been documented which shows drawings of animals that look much like what we would expect Zaglossus hacketti to look like, further indicating that this large monotreme was known to the aboriginal people.
The largest prehistoric platypus is Obdurodon tharalkooschild
.jpg)
Within the distinctive platypus beak, the animal had rows of sharp teeth, which scientists believe they used to slice and chew crayfish, frogs and small turtles.
However, for such prey this deadly threat has passed – and will never interact with river-wading humans" ankles – since the ancient platypus is extinct.
The tooth, found embedded in limestone, revealed an entirely unknown branch of the platypus"s evolution. Until now, the idiosyncratic mammal, which lays eggs and is venomous, was considered to have a single direct evolutionary lineage.
Thе platypuses were present in South America until 61m years ago, as well as Antarctica, when the two continents were joined to Australia"s land mass.
Marsupials (Marsupialia)

1. Nambaroo 2. Chaeropus 3. Zygomaturus 4. Procoptodon 5. Diprotodon
The relationships among the three extant divisions of mammals (monotremes, marsupials, and placentals) were long a matter of debate among taxonomists.Most morphological evidence comparing traits such as number and arrangement of teeth and structure of the reproductive and waste elimination systems as well as most genetic and molecular evidence favors a closer evolutionary relationship between the marsupials and placental mammals than either has with the monotremes.
The ancestors of marsupials, part of a larger group called metatherians, probably split from those of placental mammals (eutherians) during the mid-Jurassic period, though no fossil evidence of metatherians themselves are known from this time.From DNA and protein analyses, the time of divergence of the two lineages has been estimated to be around 100 to 120 mya.Fossil metatherians are distinguished from eutherians by the form of their teeth; metatherians possess four pairs of molar teeth in each jaw, whereas eutherian mammals (including true placentals) never have more than three pairs.Using this criterion, the earliest known metatherian is Sinodelphys szalayi, which lived in China around 125 mya.This makes it a contemporary to some early eutherian species which have been found in the same area.While placental fossils dominate in Asia, marsupial fossils occur in larger numbers in North America.
The oldest metatherian fossils are found in present-day China.About 100 mya, the supercontinent Pangaea was in the process of splitting into the northern continent Laurasia and the southern continent Gondwana, with what would become China and Australia already separated by the Tethys Ocean. From there, metatherians spread westward into modern North America (still attached to Eurasia), where the earliest true marsupials are found. Marsupials are difficult to distinguish from other fossils, as they are characterized by aspects of the reproductive system which do not normally fossilize (including pouches) and by subtle changes in the bone and tooth structure that show a metatherian is part of the marsupial crown group (the most exclusive group that contains all living marsupials).
The earliest definite marsupial fossil belongs to the species Peradectes minor, from the Paleocene of Montana, dated to about 65 million years ago.From their point of origin in Laurasia, marsupials spread to South America, which was possibly connected to North America at around 65 mya through a ridge that has since moved on to become the Caribbean Archipelago.Laurasian marsupials eventually died off, for not entirely clear reasons; convention has it that they disappeared due to competition with placentals, but this is no longer accepted to be the primary reason.
Marsupials, and the related Herpetotheriidae are nested within a clade of metatherians that also included a variety of Cretaceous North American taxa.
In South America, the opossums evolved and developed a strong presence, and the Paleogene also saw the evolution of shrew opossums (Paucituberculata) alongside non-marsupial metatherian predators such as the borhyaenids and the saber-toothed Thylacosmilus. South American niches for mammalian carnivores were dominated by these marsupial and sparassodont metatherians. While placental predators were absent, the metatherians did have to contend with avian (terror bird) and terrestrial crocodylomorph competition. South America and Antarctica remained connected until 35 mya, as shown by the unique fossils found there. North and South America were disconnected until about three million years ago, when the Isthmus of Panama formed. This led to the Great American Interchange. Sparassodonts disappeared for unclear reasons – again, this has classically assumed as competition from carnivoran placentals, but the last sparassodonts co-existed with a few small carnivorans like procyonids and canines, and disappeared long before the arrival of macropredatory forms like felines,while didelphimorphs (opossums) invaded Central America, with the Virginia opossum reaching as far north as Canada.

1. Nambaroo 2. Chaeropus 3. Zygomaturus 4. Procoptodon 5. Diprotodon
Marsupials reached Australia via Antarctica about 50 mya, shortly after Australia had split off. This suggests a single dispersion event of just one species, most likely a relative to South America"s monito del monte (a microbiothere, the only New World australidelphian). This progenitor may have rafted across the widening, but still narrow, gap between Australia and Antarctica. In Australia, they radiated into the wide variety seen today. Modern marsupials appear to have reached the islands of New Guinea and Sulawesi relatively recently via Australia.A 2010 analysis of retroposon insertion sites in the nuclear DNA of a variety of marsupials has confirmed all living marsupials have South American ancestors. The branching sequence of marsupial orders indicated by the study puts Didelphimorphia in the most basal position, followed by Paucituberculata, then Microbiotheria, and ending with the radiation of Australian marsupials. This indicates that Australidelphia arose in South America, and reached Australia after Microbiotheria split off.
In Australia, terrestrial placental mammals disappeared early in the Cenozoic (their most recent known fossils being 55 million-year-old teeth resembling those of condylarths) for reasons that are not clear, allowing marsupials to dominate the Australian ecosystem.Extant native Australian terrestrial placental mammals (such as hopping mice) are relatively recent immigrants, arriving via island hopping from Southeast Asia.
Genetic analysis suggests a divergence date between the marsupials and the placentals at 160 million years ago.The ancestral number of chromosomes has been estimated to be 2n = 14. A new hypothesis suggests that South American microbiotheres resulted from a back-dispersal from eastern Gondwana due to new cranial and post-cranial marsupial fossils from the Djarthia murgonensis from the early Eocene Tingamarra Local Fauna in Australia that indicate the Djarthia murgonensis is the most plesiomorphic, the oldest unequivocal australidelphian, and may be the ancestral morphotype of the Australian marsupial radiation.
The largest extinct marsupial was Diprotodon, about 3 metres (9.8 ft) long, standing 2 metres (6.6 ft) tall and weighing up to 2,786 kg (6,142 lb).
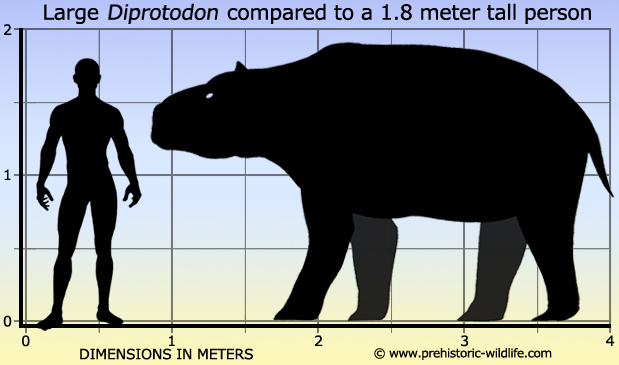
Diprotodon, meaning "two forward teeth", is the largest known marsupial ever to have lived. Along with many other members of a group of unusual species collectively called the "Australian megafauna", it existed from approximately 1.6 million years ago until extinction some 46,000 years ago (through most of the Pleistocene epoch).
Diprotodon species fossils have been found in sites across mainland Australia, including complete skulls and skeletons, as well as hair and foot impressions. Female skeletons have been found with babies located where the mother"s pouch would have been. The largest specimens were hippopotamus-sized: about 3 metres (9.8 ft) from nose to tail, standing 2 metres (6.6 ft) tall at the shoulder and weighing about 2,800 kilograms (6,200 lb). Aboriginal rock art images in Quinkan traditional country (Queensland, Australia) have been claimed to depict diprotodonts. They inhabited open forest, woodlands, and grasslands, possibly staying close to water, and eating leaves, shrubs, and some grasses.
The closest surviving relatives of Diprotodon are the wombats and the koala. It is suggested that diprotodonts may have been an inspiration for the legends of the bunyip, as some Aboriginal tribes identify Diprotodon bones as those of "bunyips".
The first recorded Diprotodon remains were discovered in a cave near Wellington in New South Wales in the early 1830s by Major Thomas Mitchell who sent them to England for study by Sir Richard Owen. In the 1840s Ludwig Leichhardt discovered many Diprotodon bones eroding from the banks of creeks in the Darling Downs of Queensland and when reporting the find to Owen commented that the remains were so well preserved he expected to find living examples in the then unexplored central regions of Australia.
The majority of fossil finds are of demographic groups indicative of diprotodonts dying in drought conditions. For example, hundreds of individuals were found in Lake Callabonna with well-preserved lower bodies but crushed and distorted heads. It is theorised several family groups sank in mud while crossing the drying lake bed. Other finds consist of age groupings of young or old animals which are first to die during a drought.
In 2012, a significant group of about 40 was found at Eulo, South-West Queensland.
Diprotodon was named by Owen (1838). It was assigned to Diprotodontidae by McKenna and Bell (1997). The historical classification of Diprotodon consisted of eight species (Diprotodon optatum Owen, 1838; Diprotodon australis Owen, 1844; D. annextans McCoy, 1861; D. minor Huxley, 1862; D. longiceps McCoy 1865; D. loderi Krefft, 1873a; D. bennettii Krefft, 1873b (nec D. bennettii Owen, 1877); and D. bennettii Owen, 1877 (nec D. bennettii Krefft, 1873b); based on size or slight morphological differences of single specimens collected from isolated geographic regions.Bimodal dental sizes, rather than a continuum of tooth sizes, and identical male and female dental morphology, indicate sexual dimorphism instead of separate species, thus providing strong evidence that the eight species are synonyms for D. optatum.
Diprotodon superficially resembled a rhinoceros without a horn. Its feet turned inwards like a wombat’s, giving it a pigeon-toed appearance. It had strong claws on the front feet and its pouch opening faced backwards. Footprints of its feet have been found showing a covering of hair which indicates it had a coat similar to a modern wombat.
Until recently it was unknown how many species of Diprotodon had existed. Eight species are described although many researchers believed these actually represented only three at most while some estimated there could be around twenty in total.
Recent research compared the variation between all of the described Diprotodon species with the variation in one of Australia’s largest living marsupials the Eastern Grey Kangaroo and found the range was comparable, with a near continent-wide distribution. This left only two possible Diprotodon species differing only in size with the smaller being around half the size of the larger. According to Gause’s "competitive exclusion principle" no two species with identical ecological requirements can coexist in a stable environment. However, both the small and large diprotodonts coexisted throughout the Pleistocene and the size difference is similar to other sexually dimorphic living marsupials. Further evidence is the battle damage common in competing males found on the larger specimens but absent from the smaller. Dental morphology also supports sexual dimorphism, with highly sexually dimorphic marsupials, such as the grey kangaroo, having different tooth sizes between males and females, but both sexes having the same dental morphology. An identical dental morphology occurs in the large and small Diprotodon. The taxonomic implication is that Owen’s original Diprotodon optatum is the only valid species.
A single sexually dimorphic species allows behavioural interpretations. All sexually dimorphic species of over 5 kilograms (11 lb) exhibit a polygynous breeding strategy. A modern example of this is the gender segregation of elephants where females and the young form family groups while lone males fight for the right to mate with all the females of the group. This behaviour is consistent with fossil finds where adult/juvenile fossil assemblages usually contain only female adult remains.
Most modern researchers including Richard Roberts and Tim Flannery argue that diprotodonts, along with a wide range of other Australian megafauna, became extinct shortly after humans arrived in Australia about 50,000 years ago.
Some older researchers including Richard Wright argue on the contrary that diprotodont remains from several sites, such as Tambar Springs and Trinkey and Lime Springs suggest that Diprotodon survived much longer, into the Holocene. Other more recent researchers, including Lesley Head and Judith Field, favour an extinction date of 28,000 - 30,000 years ago, which would mean that humans coexisted with Diprotodon for some 20,000 years.However, opponents of "late extinction" theories have interpreted such late dates based on indirect dating methods as artifacts resulting from redeposition of skeletal material into more recent strata, and recent direct dating results obtained with new technologies have tended to confirm this interpretation.
Three theories have been advanced to explain the mass extinction.
Diprotodon skull, clearly showing the large front teeth for which the genus is named and the dentition adapted for browsingAustralia has undergone a very long process of gradual aridification since it split off from Gondwanaland about 40 million years ago. From time to time the process reversed for a period, but overall the trend has been strongly toward lower rainfall. The recent ice ages produced no significant glaciation in mainland Australia but long periods of cold and very dry weather. This dry weather during the last ice age may have killed off all the large diprotodonts.
Critics point out a number of problems with this theory. First, large diprotodonts had already survived a long series of similar ice ages, and there does not seem to be any particular reason the most recent one should have achieved what all the previous ice ages had failed to do. Also, climate change apparently peaked 25,000 years after the extinctions. Finally, even during climatic extremes, some parts of the continent always remain relatively exempt: for example, the tropical north stays fairly warm and wet in all climatic circumstances; alpine valleys are less affected by drought, and so on.
The "blitzkrieg theory" is that human hunters killed and ate the diprotodonts, causing their extinction. The extinctions appear to have coincided with the arrival of humans on the continent, and in broad terms, Diprotodon was the largest and least well-defended species that died out. Similar hunting-out happened with the megafauna of New Zealand, Madagascar and many smaller islands around the world (such as New Caledonia, Cyprus, Crete and Wrangel Island), and at least in part, in the Americas—probably within a thousand years or so. Recent finds of Diprotodon bones which appear to display butchering marks lend support to this theory. Critics of this theory regard it as simplistic, arguing that (unlike New Zealand and America) there is little direct evidence of hunting, and that the dates on which the theory rests are too uncertain to be relied on. However, the high-resolution chronology of the changes supports the hypothesis that human hunting alone eliminated the megafauna.
The third theory says that humans indirectly caused the extinction of diprotodonts, by destroying the ecosystem on which they depended. In particular, early Aborigines are thought to have been fire-stick farmers using fire regularly and persistently to drive game, open up dense thickets of vegetation, and create fresh green regrowth for both humans and game animals to eat. Evidence for the fire hypothesis is the sudden increase in widespread ash deposits at the time that people arrived in Australia, as well as land-management and hunting practices of modern Aboriginal people as recorded by the earliest European settlers before Aboriginal society was devastated by European contact and disease. Evidence against the hypothesis is the fact that humans appear to have eliminated the megafauna of Tasmania without using fire to modify the environment there.
The above hypotheses are not necessarily mutually exclusive. Each of proposed mechanisms can potentially support the other two. For example, while burning an area of fairly thick forest and thus turning it into a more open, grassy environment might reduce the viability of a large browser (an animal that eats leaves and shoots rather than grasses), the reverse could also be true: removing the browsing animals (by eating them, or by any other means) within a few years produces a very thick undergrowth which, when a fire eventually starts through natural causes (as fires tend to do every few hundred years), burns with greater than usual ferocity. The burnt-out area is then repopulated with a greater proportion of fire-loving plant species (notably eucalypts, some acacias, and most of the native grasses) which are unsuitable habitat for most browsing animals. Either way, the trend is toward the modern Australian environment of highly flammable open sclerophyllous forests, woodlands and grasslands, none of which are suitable for large, slow-moving browsing animals—and either way, the changed microclimate produces substantially less rainfall.
An examination of swamp sediment cores spanning the last 130,000 years from Lynch"s Crater in Queensland suggests that hunting may have been the primary cause of the extinction. Analysis of Sporormiella fungal spores (which derive mainly from the dung of megaherbivores) in the cores shows that the megafauna of that region virtually disappeared about 41,000 years ago, at a time when climate changes were minimal; the change was accompanied by an increase in charcoal, and was followed by a transition from rainforest to fire-tolerant sclerophyll vegetation. The high-resolution chronology of the changes indicates that fire increased about a century after the disappearance of browsing megafauna, probably due to accumulation of fuel. Grass increased over the next several centuries; sclerophyll vegetation increased following a lag of another century, and a sclerophyll forest developed about a thousand years later. Earlier increases in sclerophyll vegetation during shifts to cooler, drier conditions about 120,000 and 75,000 years ago did not have any obvious impact on megafaunal abundance.
Тагове:
Encyclopedia Largest Prehistoric Animals...
Encyclopedia Largest Prehistoric Animals...
