

Прочетен: 8397 Коментари: 0 Гласове:
Последна промяна: 08.08.2021 09:34
Rhinocerotidae
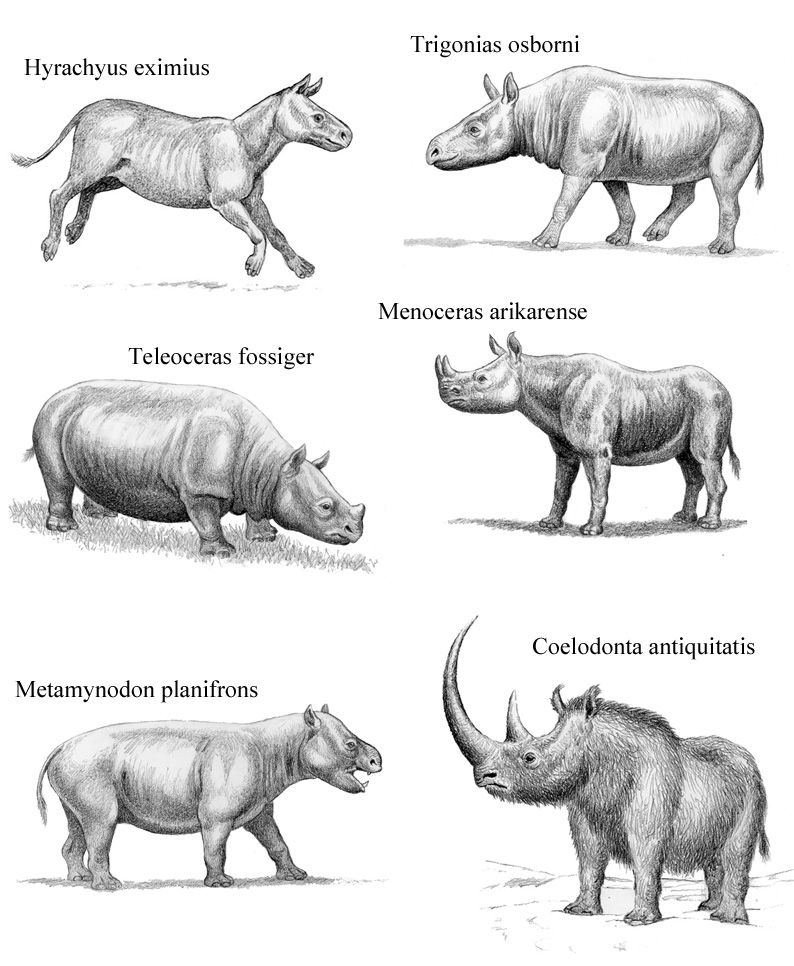
True rhinos are members of the family Rhinocerotidae. Compared to other perissodactyl families, true rhinos got off to a late start and do not appear in the fossil record until about 40 million years ago; most of the other perissodactyl families appear 55.5-46 million years ago. We think of rhinos as being characterized by a nasal horn, but in fact most members of the group are hornless. True rhinos are united by having large, tusk-like lower incisor teeth that sharpen on smaller chisel-shaped upper incisors. Curiously, the living African rhinos have lost their incisors, although they have many other anatomical features that indicate they are closely related to other true rhinos.
The earliest members of Rhinocerotidae were small and numerous; at least 26 genera lived in Eurasia and North America until a wave of extinctions in the middle Oligocene wiped out most of the smaller species. However, several independent lineages survived. Menoceras, a pig-sized rhinoceros, had two horns side-by-side. The North American Teleoceras had short legs, a barrel chest and lived until about 5 million years ago. The last rhinos in the Americas became extinct during the Pliocene. Modern rhinos are thought to have begun dispersal from Asia during the Miocene. Two species survived the most recent period of glaciation and inhabited Europe as recently as 10,000 years ago: the woolly rhinoceros and Elasmotherium.

The woolly rhinoceros appeared in China around 1 million years ago and first arrived in Europe around 600,000 years ago. It reappeared 200,000 years ago, alongside the woolly mammoth, and became numerous. Elasmotherium was two meters tall, five meters long and weighed around five tons, with a single enormous horn, hypsodont teeth and long legs for running. Both the Coelodonta and Elasmotherium survived through the late Pleistocene when climate fluctuations, increased predation by newer predators such as hyenas and human hunting led to their extinction. There may have been a remnant population of Elasmotherium, however, in the south of Western Siberia (the area that is today Kazakhstan) as recently as 29,000 years ago.
Of the extant rhinoceros species, the Sumatran rhino is the most archaic, first emerging more than 15 million years ago. The Sumatran rhino was closely related to the woolly rhinoceros, but not to the other modern species. The Indian and Javan rhinos are closely related and form a more recent lineage of Asian rhino. The ancestors of early Indian and Javan rhino diverged 2–4 million years ago.
The origin of the two living African rhinos can be traced to the late Miocene (6 mya) species Ceratotherium neumayri. The lineages containing the living species diverged by the early Pliocene (1.5 mya), when Diceros praecox, the likely ancestor of the black rhinoceros, appears in the fossil record. The black and white rhinoceros remain so closely related that they can still mate and successfully produce offspring.
Aphelops Mutilus

Aphelops is an extinct genus of rhinoceros endemic to North America during the Miocene through the Pliocene, living from 20.43—5.330 mya, existing for approximately 15.1 million years.
Aphelops Mutilus was a rhinoceros, an ancestor of the white rhino and black, but also the woolly rhinoceros. Easily exceeded 4.2 meters long (14 ft.) and was close to 3 meters tall (9 ft,). They were immense.
A large male white rhino can exceed 3,000 kg (6.600 lb), but easily surpassed Aphelops 4 tons (8.800 lb). Its constitution was robust, the larger body and smaller legs. Looked more like a hippo rhino.
Petite legs, big, round and with a huge rib cage body, without much hump like a rhinoceros. In fact, after analyzing the skeleton and teeth, experts concluded that the Aphelops Mutilus spent much of their time in the water, as hiccups live. Once out, he would be helpless, despite its large size, face formidable predators as Hyaenodon or Entelodon (or giant boar). If only I had a horn to defend ...
This large herbivore existed in the Miocene, between 20 and 5 million behind in what is now North America. Besides Aphelops Mutilus have been classified two species: Aphelops malacorhinus and Aphelops megalodus.
Chilotherium is an extinctgenus of rhinoceros endemic to Eurasia during the Miocene through Pliocene living for 13.7—3.4 mya, existing for approximately 10.3 million years. It was a large, robust animal reaching 1.5-1.8 m in height and a weight between 1 and 2.5 tons, depending on the species
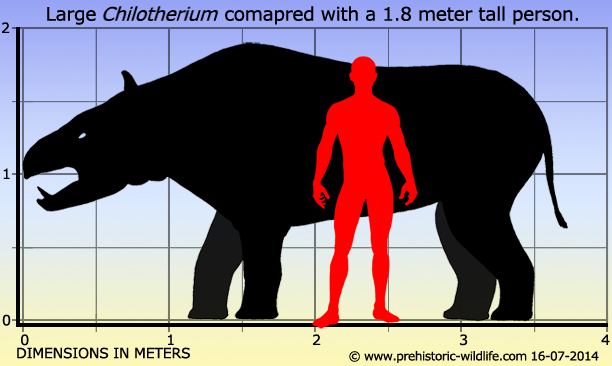
Chilotherium is a genus of prehistoric rhinoceros that seems to have had a geographic distribution spreading across Asia all the way up to Eastern Europe. Chilotherium had no nasal horns like rhinoceros are often portrayed as having, but Chilotherium still remained quite unique. Two tusks formed from enlarged incisor teeth rose up from the lower jaw, and while these tusks were present in male and female Chilotherium, they seem to have been larger in the males. This would indicate that the upward facing tusks had a display purpose, thought he fact that they were also present in females would suggest a species recognition purpose as well as a possible practical application. Some species of Chilotherium are noted as browsers, while others seem to be dedicated grazers.
A fossil skull of a female Chilotherium has been preserved with partially healed tooth marks that may have been caused by an attacking Dinocrocuta.
Stephanorhinus kirchbergensis
Stephanorhinus is an extinct genus of rhinoceros native to northern Eurasia that lived during the middle and early late Pleistocene epoch. It had two horns and was a relatively large rhino. It weighted over 3,000 kg and measured about 1.80 - 2 m tall and 3.20 - 4 m in length, having a size similar to a white rhino. In the late Pleistocene, there were two species of Stephanorhinus in Eurasia, the large Merck"s rhinoceros Stephanorhinus kirchbergensis and the narrow-nosed rhinoceros Stephanorhinus hemiotechus. Both species may have evolved from Stepahnorhinus etruscus. The narrow-nosed rhinoceros appeared in Europe in the early middle Pleistocene. It is known here from many localities between, Spain, Italy, Germany and the British Islands. Apart from Europe it is also known from Syria, Israel, the Caucasus and from one late Pleistocene locality close to Lake Baikal.
Coelodonta
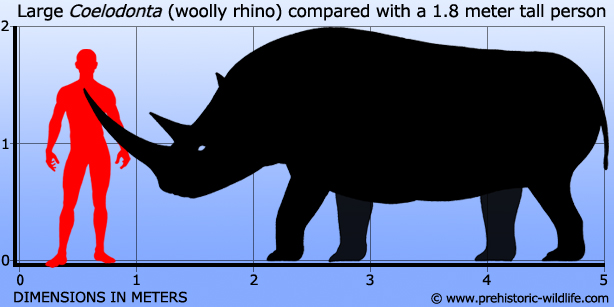
Coelodonta is one of the most commonly represented ‘ice age’ mammals, yet surprisingly it often only gets a name mention. Popularly known as the woolly rhino, Coelodonta resembled the large rhinos that we know today from Africa, but with a complete covering of fur over its body. This was the main survival adaptation of Coelodonta which inhabited most of Eurasia for over two and a half million years.
Coelodonta lived at a time that saw a series of glaciations across the Northern hemisphere that saw sheets of ice sweeping over much of the land, to receding back before covering the land again. This toing and froing of the ice sheets, combined with the colder climate that caused much of the lower soil depths to be permanently frozen, created vast expanses of frozen plains that were covered in grasses interspersed with low growing vegetation, and it is this ecosystem that Coelodonta seems to have been most adapted to. As with many similar herbivores, Coelodonta had sharp incising teeth at the front of the mouth and mashing molar teeth at the back. Between these two sets of teeth was a gap called the diastema, something else that is common amongst herbivorous mammals.
It’s uncertain exactly what kind of herbivore Coelodonta was. Some people think that Coelodonta was a grazer that cropped the grass plains like a cow, while others believe that it was a browser than fed from low growing plants. Either one is plausible; although most lean towards the grazing idea as grasses would have been much more abundant than more complex low growing plants. Coelodonta is thought to have used a fermentation method of processing the cellulose rich grasses in order to get the full nutritional benefit from the nutritionally poor vegetation of the ecosystem. This is actually a very clever method of digestion to adopt since as the grass is broken down by fermentation inside the gut of Coelodonta, it generates a small amount of heat that would have the added effect of warming the body from the inside.
This method of digestion seems to have been very efficient for Coelodonta as specimens where the main body is still preserved show that Coelodonta had a hump that rose up from its back above the shoulder blades. This hump was supported from within by the forwards dorsal vertebrae that had elongated neural spines growing from them, much larger the neural spines of the other vertebrae. This hump would have served as fat storage so that Coelodonta could build up fat reserves in the milder spring and summer so that it could better survive the colder winter when the plants had begun to die back, and possibly even had a deep covering of snow and ice.
Efficient processing of plants that have a low nutritional value is important for an animal like Coelodonta, but just as important for survival is efficient energy use and conservation. As a mammal, Coelodonta was certainly warm blooded, which means that the body works to try and maintain a stable temperature different from it environment (in this case much higher). Usually this is done by a process of involuntary muscle actions to generate heat such as the rapid and repeated constriction of muscle fibres, better known as ‘shivering’. In the short term shivering is not a problem, but prolonged shivering results in more calories being burned (used up), and the more that are burned the more likely that an animal will use up all it has. With no more calories to use, and not enough coming from the plants to maintain the level of use, an animal will stop shivering and quickly succumb to the cold, quite possibly dying from exposure.
As briefly mentioned above, Coelodonta had a covering of fur over its body, something which led to the name ‘woolly rhino’. This was the main line of defence against the cold, and would have trapped layers of air near to the skin. With these layers protected from mixing with the outer air they would be warmed by the body, and because this layer of inner air did not require any energy to maintain, it would have been like a blanket that slowed down the rate of heat loss from the body to the outside environment. A similar effect to this is simply wearing clothes as these layers of fabric trap pockets of air against your body to keep you warm. The effect is especially pronounced for thicker garments such as jumpers which offer a greater level of insulation. In addition to the fur, the body proportions of Coelodonta also helped to prevent heat loss. For example, the short legs of Coelodonta are a further adaption to the cold environment as they would reduce the surface area exposed to the cold climate, reducing the area for heat loss to take place.
No description of a rhino would be complete without mentioning the horn and this goes double for Coelodonta as it had two pronounced horns rising from its snout. The front horn at up to two meters long was the longer of the two, with the second horn rising from the middle of the snout being just over half to two thirds as big. The classical explanation for these horns is that they were what are termed sexually selected characteristics. This is based upon the knowledge that the horn would have been growing throughout the animals life, no more than a stump in a juvenile to fully developed in mature individuals. An older animal would have a more developed horn than younger individuals signalling to members of the opposite sex that it had the genetic makeup and success to make it to later life, and was more deserving of passing its genes down to the next generation than lesser individuals that had less developed horns. Such reasoning would explain the progression to larger horn sizes.
However there is a second theory regarding the front horn that is both an alternative and possible addition to the above theory. The front horn is strongly curved so that it extends out beyond the end of the snout. This horn is also laterally compressed so that when viewed from the front the horn looks more like a blade rather than a cone like in other genera. This leads to the popular interpretation the front horn was not just a display device but an actual tool that Coelodonta used to scrape snow off the ground as it moved its head from side to side. This would expose buried grasses that allowed Coelodonta to feed further without using energy to walk to an area that was uncovered, and would have been of particular use when Coelodonta was in areas that had frequent snowfall, but not a permanent covering.
The earliest remains of Coelodonta are from India and have been dated back to the end of the Pliocene period. The majority of other Coelodonta remains so far known are from Europe and Russia and these date back to the Calabrian of the Pleistocene, which suggests that Coelodonta first emerged in central Asia and then expanded its range. This expansion could have been synchronised to the availability of tundra-like environments that were constantly changed from the varying expansion and receding of the ice sheets that once covered the northern hemisphere.
One of the best examples of Coelodonta comes from a Tar Pit in Poland (near Starunia) that had its body frozen and preserved. Before this time the only visual representation of the living Coelodonta was in the form of cave art that had been made by ancient human beings.
Encyclopedia Largest prehistoric animals...
Encyclopedia Largest prehistoric animals...
