
Постинг
09.01.2016 01:16 -
Encyclopedia Largest Prehistoric Animals Vol.1 Vertebrates part 3 Dinosaurs ch.2 Sauropods - Who"s The Biggest Sauropod
Автор: valentint
Категория: Забавление

Прочетен: 4361 Коментари: 0 Гласове:
Последна промяна: 08.06.2020 17:42
Прочетен: 4361 Коментари: 0 Гласове:
0
Последна промяна: 08.06.2020 17:42
Sauropods (Sauropoda)
Amphicoelias,the largest land animal vs possible heaviest Bruhathkayosaurus
From left to right :Camarasaurus lentus, Giraffatitan brancai, Diplodocus hallorum, Spinophorosaurus nigerensis, Glacialisaurus hammeri, Dicraeosaurus hansemanni, Apatosaurus louisae, Daxiatitan binglingi, Brachiosaurus altithorax, Lamplughsaura dharmaramensis, Supersaurus vivianae, Argyrosaurus superbus, Diplodocus carnegii, Apatosaurus ajax, Mamenchisaurus sinocanadorum, Atlasaurus Imelakei, Cetiosaurus oxoniensis, Omeisaurus junghsiensis, Sauroposeidon proteles, Barosaurus lentus,Lusotitan atalaiensis, Amphicoelias altus, Ampelosaurus atacis, Ruyangosaurus giganteus, Paralititan stromeri, Turiasaurus riodevensis, Jobaria tiguidensis, Alamosaurus sanjuanensis, Antarctic titanosaur, Vulcanodon karibaensis, Abydosaurus mcintoshi, Notocolossus gonzalezparejasi, Nigersaurus taqueti, Rhoetosaurus brownei, Brontosaurus parvus, Puertasaurus reuili, Patagotitan mayorum, Diamantinasaurus matildae, Saltasaurus loricatus, Futalognkosaurus dukei, Huanghetitan ruyangensis, Barapasaurus tagorei, Antarctosaurus giganteus, Magyarosaurus dacus, Austrosaurus, Malawisaurus, Savannasaurus elliottorum, Argentinosaurus huinculensis, Dreadnoughtus schrani, Isisaurus colberti and Amargasaurus cazaui.
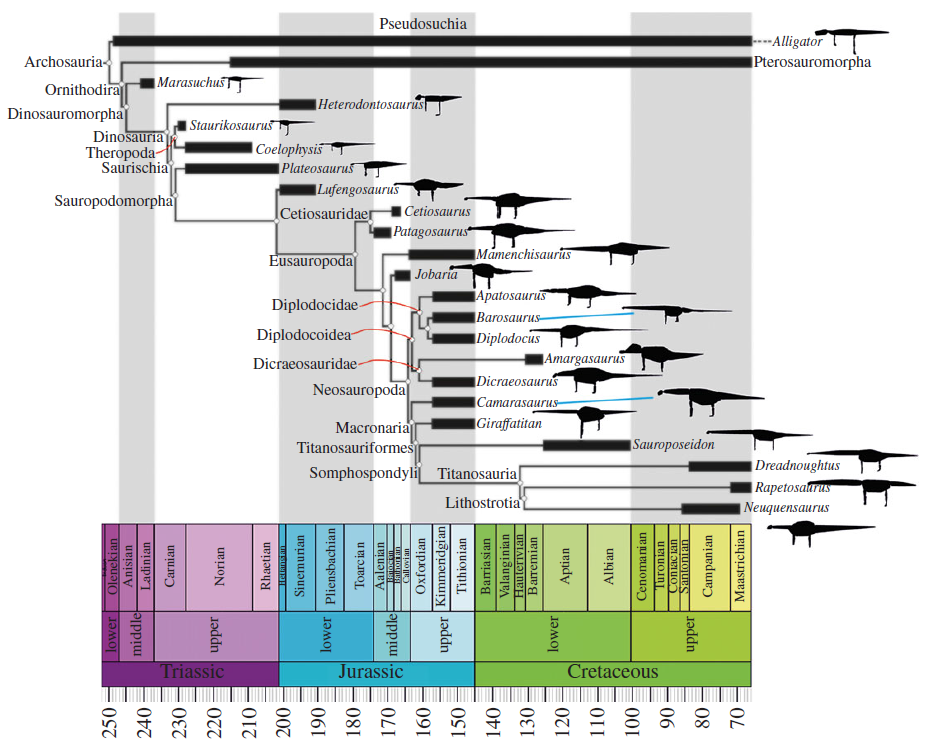
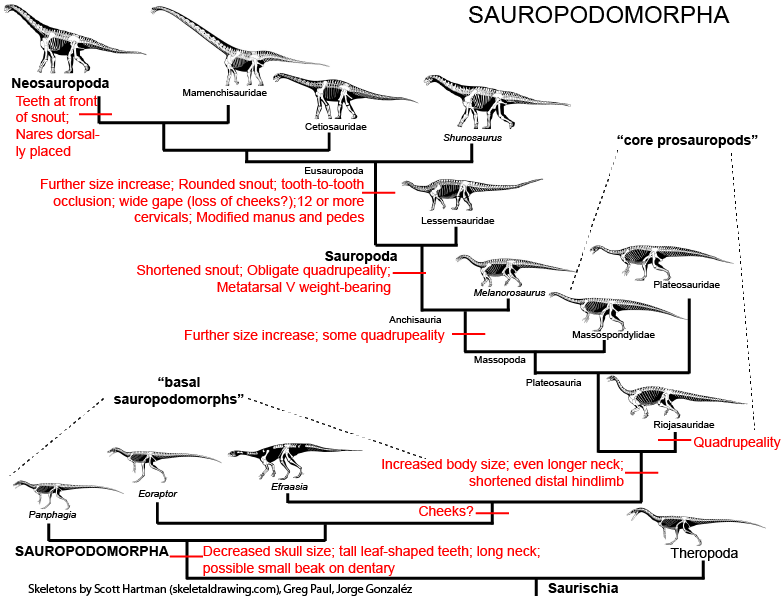
The sauropods are the gigantic long-necked plant-eating dinosaurs, and include such well-known names as Apatosaurus ("Brontosaurus"), Brachiosaurus, Cetiosaurus, and Diplodocus, as well as numerous other species and genera. They were the largest of all land animals, and among the most successful and long-lived of the dinosaurs.
Among the very first dinosaurs to evolve in the Late Triassic Period, about 230 million years ago (Mya), they became the dominant herbivores by halfway through the late Triassic (during the Norian stage). Their perceived decline in the early Cretaceous is most likely a bias in fossil sampling, as most fossils are known from Europe and North America. Sauropods were still the dominant herbivores in the Gondwanan landmasses, however. The spread of flowering plants (angiosperms) and "advanced" ornithischians, another major group of herbivorous dinosaurs (noted for their highly developed chewing mechanisms), are most likely not a major factor in sauropod decline in the northern continents.Like all non-avian dinosaurs, the sauropodomorphs became extinct 66 Mya, during the Cretaceous–Paleogene extinction event.

The earliest and most basal sauropodomorphs known are Chromogisaurus novasi and Panphagia protos, both from the Ischigualasto Formation, dated to 231.4 million years ago (late Ladinian age of the Middle Triassic according to the ICS; alternately called the early Carnian age of the Late Triassic in the system used by the Geological Society of America).Some studies have found Eoraptor lunensis (also from the Ischigualasto Formation), traditionally considered a theropod, to be an early member of the sauropodomorph lineage, which would make it the most basal sauropodomorph known.
The earliest known species, the small Vulcanodon (6 1/2 metres) dates back to the Sinemurian epoch, while the latest include a number of species such as the small Titanosaurus indicus and Hypselosaurus priscus (lengths about 12 metres), and the large Alamosaurus sanjuanensis (20 metres), were still flourishing right up until the terminal Cretaceous extinction. It was at one time thought on the basis of the North American fossil record) that the sauropods became rare after the Late Jurassic, but in most parts of the world they continued as a common element of the megafauna until the very end of the Cretaceous. Throughout their long history they continued to evolve and branch out new species and families. There are at least three peaks of sauropod diversity in the Late Jurassic, late Early Cretaceous and latest Cretaceous, corresponding to high sea levels and increased speciation resulting from geographical isolation.

Sauropods were long believed to be semi-aquatic swamp wallowers, relying on the bouyancy of water to support their massive bodies. But analysis of their skeletons, in comparison with those of large terrestrial and semi-aquatic animals, and of sedimentation where their fossils have been found, show that sauropods were fully terrestrial:
Not only were sauropods as terrestrial as elephants, but fossil trackways indicate that they lived in herds, again like elephants today. It must have been a truely awesome sight to watch a herd of brontosaurs crossing a Mesozoic floodplain; evoking the same sense of awe, and puniness in one"s own being in comparison, as one would feel when observing whales close up.
The Sauropoda include the largest animals ever to walk on land. These gigantic herbivores reaches lengths of 15 to 25 metres or more (the very largest may have reached 60 metres) and weights of 15 to 30 or even 80 or 150 tonnes.
There are currently eight valid families of sauropods: Vulcanodontidae, Cetiosauridae, Brachiosauridae, Camarasauridae, Diplodocidae, Euhelopodidae, Dicraeosauridae and Titanosauridae.
However, this is certainly an underdestimate of the true diversity of these amazing beasts. Ironically, despite their great size (which would one would think would enable more frequent preservation of remains) the fossil record is quite poor, perhaps due to the large skeletons being skattered before sediment settles to cover them. Most sauropod remains are actually scattered and isolated bones and teeth.
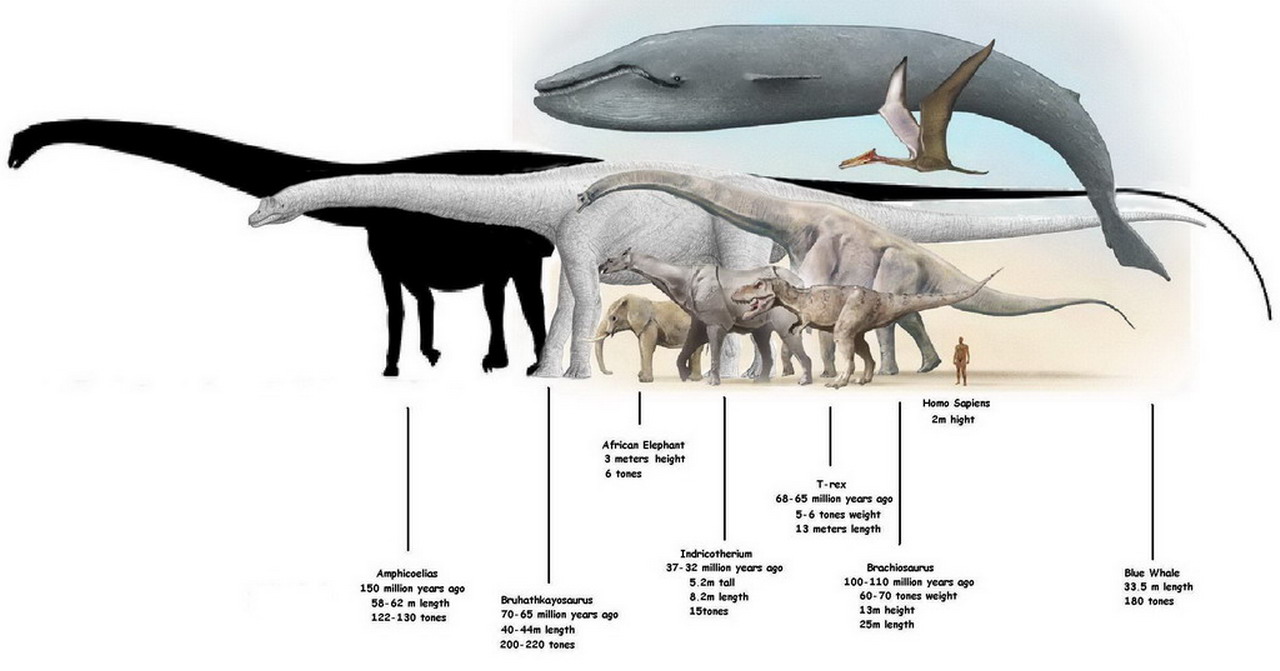
The Sauropods" most defining characteristic was their size. Even the smallest of the species, the dwarf sauropods, grew up to 5 to 6 metres in length, and were counted as some of the largest dinosaurs in their ecosystem. As the largest animals to have ever walked on land, the only competitor in the history of the animal kingdom are the rorqual whales, such as the blue whale.
 While many dinosaurs of different genus had individual variations from their related cousins, the sauropods had little variation between the individual species, possibly due to the forced shared evolution caused by size constraints. However, some members of the genus had individual characteristics, such as the diplodocids, who had extrememly long, whip-like tails, possibly as a defense mechanism against predators. Seismosaurus,the longest of the sauropods, would reach lengths of around 39–52 metres, most likely around 45 metres. Other sauropods, such as the brachiosaurids, were extremely tall, and had high shoulders and extremely long necks used for foraging in the high tree canopies. The Sauroposeidon was most likely the tallest of the sauropods, reaching a height of 18 metres. In terms of today"s animal kingdom, the tallest living animal is the giraffe, which reaches only 4.8 to 5.5 metres in height. Due to their massive height and length, the sauropods were also the largest dinosaurs to walk on the earth. One of heaviest of the class was Argentinosaurus, which most likely weighed approximately 75 to 100 metric tonnes. Other sauropods were of comparable size to the Argentinosaurus, but there is some circumstantial evidence that an even more massive titanosaurian, the Bruhathkayosaurus, weighed a possible 175 to 220 tonnes, although a land animal this size would not have been able to support it"s own weight. By comparison, the largest land animal alive today is the Savannah elephant, which weighs only 10 tonnes.
While many dinosaurs of different genus had individual variations from their related cousins, the sauropods had little variation between the individual species, possibly due to the forced shared evolution caused by size constraints. However, some members of the genus had individual characteristics, such as the diplodocids, who had extrememly long, whip-like tails, possibly as a defense mechanism against predators. Seismosaurus,the longest of the sauropods, would reach lengths of around 39–52 metres, most likely around 45 metres. Other sauropods, such as the brachiosaurids, were extremely tall, and had high shoulders and extremely long necks used for foraging in the high tree canopies. The Sauroposeidon was most likely the tallest of the sauropods, reaching a height of 18 metres. In terms of today"s animal kingdom, the tallest living animal is the giraffe, which reaches only 4.8 to 5.5 metres in height. Due to their massive height and length, the sauropods were also the largest dinosaurs to walk on the earth. One of heaviest of the class was Argentinosaurus, which most likely weighed approximately 75 to 100 metric tonnes. Other sauropods were of comparable size to the Argentinosaurus, but there is some circumstantial evidence that an even more massive titanosaurian, the Bruhathkayosaurus, weighed a possible 175 to 220 tonnes, although a land animal this size would not have been able to support it"s own weight. By comparison, the largest land animal alive today is the Savannah elephant, which weighs only 10 tonnes.
The sauropods were herbivorous creatures, and usually were quadrupeds due to their massive girth. They shared a common characteristic of small heads, with large, massive bodies and long, counter-balancing tails. At least some of the species were egg-layers, as evidence has been found that the camarasaurs and titanosaurs laid eggs. Due to their gigantic size, sauropods had thick legs to support their weight, with blunt, balanced feet that ended in five toes.
Amphicoelias fragillimus is a genus of herbivious sauropoddinosaur and is the largest animal to have ever live, bigger than the blue whale.

A mega-sauropod, Amphicoelias fragillimus, is a contender for the largest dinosaur in history. It has been estimated at 58 metres (190 ft) in length and 122,400 kilograms (269,800 lb) in weight. Unfortunately, the fossil remains of this dinosaur have been lost. Amphicoelias (meaning "biconcave", from the Greek amphi: "on both sides", and koilos: "hollow, concave") is a genus of herbivorous sauropod dinosaur that is probably synonymous with the genus Diplodocus. It includes what has sometimes been estimated to be the largest dinosaur specimen ever discovered, originally named "A. fragillimus". Based on surviving descriptions of a single fossil bone, scientists had over the years estimated A. fragillimus to have been the longest known vertebrate at 58 metres (190 ft) in length, with potentially a mass of up to 122.4 tonnes (135 short tons). However, because the only fossil remains were lost at some point after being studied and described in the 1870s, evidence survived only in drawings and field notes. More recent analysis of the surviving evidence, and the biological plausibility of such a large land animal, has suggested that the enormous size of this animal were over-estimates due partly to typographical errors in the original 1878 description.
The type species of Amphicoelias, A. altus, was named by paleontologist Edward Drinker Cope in December 1877 (though not published until 1878) for an incomplete skeleton consisting of two vertebrae, a pubis (hip bone), and a femur (upper leg bone).Cope also named a second species, A. fragillimus, in the same paper. However, all subsequent researchers have considered A. fragillimus to be a synonym of A. altus. Even by 1881 however, it was recognized that A. altus could not be distinguished from other genera, as the features described by Cope were misinterpreted and are widespread.In 1921, Osborn and Mook assigned additional bones to A. altus—a scapula (shoulder blade), a coracoid (shoulder bone), an ulna (lower arm bone), and a tooth. Henry Fairfield Osborn and Charles Craig Mook noted the overall close similarity between Amphicoelias and Diplodocus, as well as a few key differences, such as proportionally longer forelimbs in Amphicoelias than in Diplodocus. The dentition of Amphicoelias is homodont. Its teeth are shaped like long slender cylindrical rods, are spaced apart and project forward towards the front of the mouth. The femur of Amphicoelias is unusually long, slender, and round in cross section; while this roundness was once thought to be another distinguishing characteristic of Amphicoelias, it has since been found in some specimens of Diplodocus as well. A. altus was also similar in size to Diplodocus, estimated to be about 25 m (82 ft) long.While most scientists have used these details to distinguish Amphicoelias and Diplodocus as separate genera, at least one has suggested that Amphicoelias is probably the senior synonym of Diplodocus.
Amphicoelias fragillimus was collected by Oramel Lucas, a fossil collector employed by E. D. Cope, shortly after he was hired by Cope in 1877. Lucas discovered a partial vertebra (the neural arch and spine) of the new sauropod species in Garden Park, north of Caсon City, Colorado, close to the quarry that yielded Camarasaurus. The vertebra was in poor condition, but astonishingly large, measuring 1.5 metres (4.9 ft) up to 2.7 metres (8.9 ft) in height.Lucas shipped the specimen to Cope in the spring or early summer of 1878, and Cope published it as the holotype specimen (catalogue number AMNH 5777) of a new species, A. fragillimus, that August. The name derives from the Latin fragillimus ("very fragile"), referring to the delicateness of the bone produced by very thin laminae (vertebral walls). As revealed in Cope"s notebooks, which he recorded based on Lucas" report on excavation site locations in 1879, the specimen came from a hill south of the Camarasaurus quarry now known as "Cope"s Nipple." While Cope originally wrote that the site belonged to the Dakota Formation (mid-Cretaceous in age), the presence of dinosaurs such as Camarasaurus in the same rocks indicates that they probably belong to the Morrison Formation, which places the age of the site at 150 million years ago in the late Jurassic period, specifically the Tithonian age.
The gigantic bones attributed to A. fragillimus have often been ignored in summaries of the largest dinosaurs partly because, according to various subsequent reports, the whereabouts of both the vertebra and the femur are unknown, and all attempts to locate them have failed.Carpenter, in 2006, presented a possible scenario for the disappearance of the A. fragillimus specimens. As Cope noted in his description, the neural arch bone material was very fragile, and techniques to harden and preserve fossil bone had not yet been invented (Cope"s rival, paleontologist O.C. Marsh, was the first to use such chemicals, in the early 1880s). Carpenter observed that the fossil bones known from the A. fragillimus quarry would have been preserved in deeply weathered mudstone, which tends to crumble easily and fragment into small, irregular cubes. Therefore, the bone may have crumbled badly and been discarded by someone at the American Museum of Natural History (possibly even by Cope himself) soon after he illustrated it in rear view for his paper. Carpenter suggested that this may explain why Cope drew the vertebra in only one view, rather than from multiple angles as he did for his other discoveries.
In 1994, an attempt was made to relocate the original quarry where A. fragillimus and other species had been found, using ground-penetrating radar in an attempt to image bones still buried in the ground. This attempt failed because the fossilized mudstone bones were the same density as the surrounding rock, making it impossible to differentiate the two. A study of the local topography also showed that the fossil-bearing rock strata was severely eroded, and probably was so at the time Lucas made his discovery of A. fragillimus, indicating that a majority of the skeleton was gone by the time the vertebra and femur were recovered.
Cope"s description of A. fragillimus has been met with skepticism, with some researchers noting that there were typographical errors in his measurements. For example, the measurement units are given in (obviously incorrect) centimeters rather than millimeters. Carpenter argued that there is every reason to take Cope at his word, noting that the paleontologist"s reputation was at stake. The discovery took place during the Bone Wars, and Cope"s rival Marsh, who was "ever ready to humiliate" Cope, never called the claims into question. Marsh was known to have employed spies to monitor Cope"s discoveries, and may have even had confirmation.
The enormous size of the Amphicoelias fragillimus bones. Paleontologists Henry Fairfield Osborn and C.C. Mook in 1921, as well as John S. McIntosh in 1998, also accepted Cope"s data without question in published reviews. In 2015, Woodruff and Foster published an analysis of the evidence and circumstances surrounding the publication and interpretation of A. fragillimus.
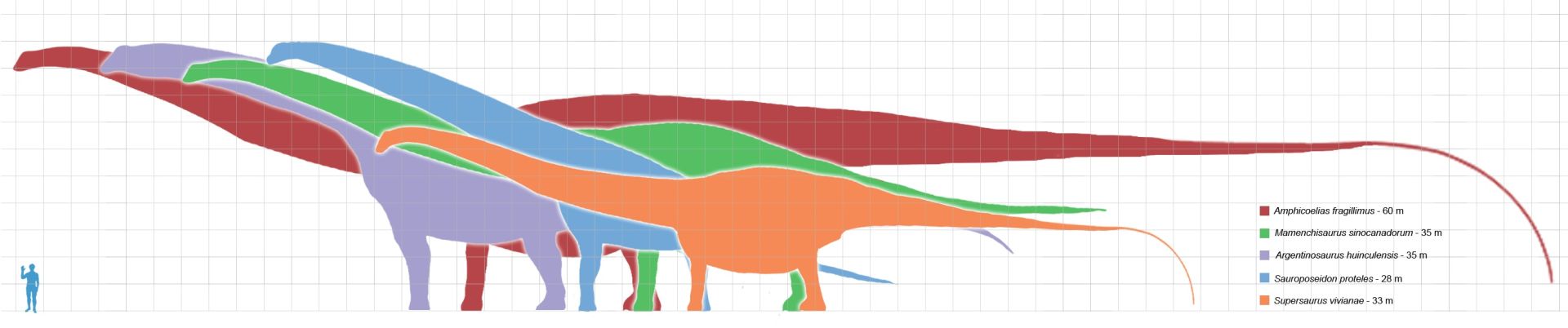
Producing an estimate of the complete size of A. fragillimus requires scaling the bones of better-known species of diplodocid (a family of extremely long and slender sauropods) in the assumption that their relative proportions were similar. In his original paper, Cope did this by speculating on the size of a hypothetical A. fragillimus femur (upper leg bone). Cope noticed that in other sauropod dinosaurs, specifically A. altus and Camarasaurus supremus, the femora were always twice as tall as the tallest dorsal vertebra, and estimated the size of an A. fragillimus femur to be 3.7 m (12 ft) tall.
In 1994, using the related Diplodocus as a reference, Gregory S. Paul estimated a femur length of 3.1 to 4 metres (10 to 13 ft) for A. fragillimus.The 2006 re-evaluation of A. fragillimus by Ken Carpenter also used Diplodocus as a scale guide, finding a femur height of 4.3 to 4.6 metres (14 to 15 ft).Carpenter went on to estimate the complete size of A. fragillimus, though he cautioned that relative proportions in diplodocids could vary from species to species. Assuming the same proportions as the well-known Diplodocus, Carpenter presented an estimated total length of 58 m (190 ft), which he noted fell within the range presented by Paul in 1994 (40 to 60 metres (130 to 200 ft)). Carpenter pointed out that even the lowest length estimates for A. fragillimus were higher than those for other giant sauropods, such as the diplodocid Supersaurus (32.5 metres (107 ft)), the brachiosaurid Sauroposeidon (34 metres (112 ft)), and the titanosaur Argentinosaurus (30 metres (98 ft)). Carpenter presented more speculative, specific proportions for A. fragillimus (again, based on a scaled-up Diplodocus), including a neck length of 16.75 metres (55.0 ft), a body length of 9.25 metres (30.3 ft), and a tail length of 32 metres (105 ft). He estimated the total forelimb height at 5.75 metres (18.9 ft) and hind limb height at 7.5 metres (25 ft), and the overall height (at the highest point on the back) at 9.25 metres (30.3 ft). By comparison the blue whale, which is on average the longest living creature, reaches 30 metres (98 ft) in length.
 While A. fragillimus was relatively thin, its enormous size still made it very massive. Weight is much more difficult to determine than length in sauropods, as the more complex equations needed are prone to greater margins of error based on smaller variations in the overall proportions of the animal. Carpenter used Paul"s 1994 estimate of the mass of Diplodocus carnegii (11.5 metric tons (11.3 long tons; 12.7 short tons)) to speculate that A. fragillimus could have weighed up to 111.0 metric tons (109.2 long tons; 122.4 short tons).The heaviest blue whale on record weighed about 190 metric tons (190 long tons; 210 short tons), and the heaviest dinosaur known from reasonably good remains, Argentinosaurus, weighed 80 to 100 metric tons (79 to 98 long tons; 88 to 110 short tons), although if the size estimates can be validated, it could still be lighter than Bruhathkayosaurus, which has been estimated to have weighed 139 metric tons (137 long tons; 153 short tons)., but is also known from highly fragmentary remains.
While A. fragillimus was relatively thin, its enormous size still made it very massive. Weight is much more difficult to determine than length in sauropods, as the more complex equations needed are prone to greater margins of error based on smaller variations in the overall proportions of the animal. Carpenter used Paul"s 1994 estimate of the mass of Diplodocus carnegii (11.5 metric tons (11.3 long tons; 12.7 short tons)) to speculate that A. fragillimus could have weighed up to 111.0 metric tons (109.2 long tons; 122.4 short tons).The heaviest blue whale on record weighed about 190 metric tons (190 long tons; 210 short tons), and the heaviest dinosaur known from reasonably good remains, Argentinosaurus, weighed 80 to 100 metric tons (79 to 98 long tons; 88 to 110 short tons), although if the size estimates can be validated, it could still be lighter than Bruhathkayosaurus, which has been estimated to have weighed 139 metric tons (137 long tons; 153 short tons)., but is also known from highly fragmentary remains.
Edward Drinker Cope described his finds in two 1878 issues of the American Naturalist, and assigned them to the new genus Amphicoelias. He placed it in a unique family, Amphicoeliidae, though this is now considered a nomen oblitum (forgotten name). The genus is usually assigned to the family Diplodocidae, though some modern analyses have found it at the base of the larger group Diplodocoidea or as a diplodocid incertae sedis (uncertain placement). The first named species in the genus, Amphicoelias altus (holotype specimen AMHD 5764), was discovered by Cope in 1877. But while it is only represented by a partial skeleton, there are enough diagnostic characteristics to provisionally define the genus. A. altus is known from better remains, but is smaller than A. fragillimus. Cope also named a second species in 1878: Amphicoelias latus.
The third named Amphicoelias species, A. fragillimus, was known only from a single, incomplete 1.5 m tall neural arch (the part of a vertebra with spines and processes), either last or second to last in the series of back vertebrae, D (dorsal) 10 or D9. Based only on an illustration published in 1878, this vertebra would have measured 2.7 metres (8.9 ft) tall in life. However, it has been argued that the scale bar in the published description contained a typographical error, and the fossil vertebra was in fact only 1.38 metres (4.5 ft) tall. In addition to this vertebra, Cope"s field notes contain an entry for an "[i]mmense distal end of femur”, located only a few tens of meters away from the giant vertebra. It is likely that this undescribed leg bone belonged to the same individual animal as the neural spine.
In 2007, John Foster suggested that the differences usually cited to differentiate Amphicoelias altus from the more well known Diplodocus are not significant and may be due to individual variation. Foster argued that Amphicoelias is probably the senior synonym of Diplodocus, and that if further research bears this out, the familiar name Diplodocus would need to be abandoned in favor of Amphicoelias, as was the case with Brontosaurus and its senior synonym Apatosaurus.In 2015, Woodruff and Foster reiterated this conclusion, stating that there is only one species of Amphicoelias and that it could be referred to Diplodocus as Diplodocus altus. They considered the name Amphicoelias to be a nomen oblitum.
In his 2006 re-evaluation, Carpenter examined the paleobiology of giant sauropods, including Amphicoelias, and addresses the question of why this group attained such a huge size. He pointed out that gigantic sizes were reached early in sauropod evolution, with very large sized species present as early as the late Triassic Period, and concluded that whatever evolutionary pressure caused large size was present from the early origins of the group. Carpenter cited several studies of giant mammalian herbivores, such as elephants and rhinoceros, which showed that larger size in plant-eating animals leads to greater efficiency in digesting food. Since larger animals have longer digestive systems, food is kept in digestion for significantly longer periods of time, allowing large animals to survive on lower-quality food sources. This is especially true of animals with a large number of "fermentation chambers" along the intestine which allow microbes to accumulate and ferment plant material, aiding digestion. Throughout their evolutionary history, sauropod dinosaurs were found primarily in semi-arid, seasonally dry environments, with a corresponding seasonal drop in the quality of food during the dry season. The environment of Amphicoelias was essentially a savanna, similar to the arid environments in which modern giant herbivores are found, supporting the idea that poor-quality food in an arid environment promotes the evolution of giant herbivores. Carpenter argued that other benefits of large size, such as relative immunity from predators, lower energy expenditure, and longer life span, are probably secondary advantages.
Bruhathkayosaurus

Bruhathkayosaurus ( meaning "huge bodied lizard") is a dinosaur that some researchers claim to have been the largest dinosaur that ever lived. The accuracy of this claim, however, has been mired in controversy and debate. All the estimates are based on Yadagiri and Ayyasami"s 1989 paper, which announced the find.
The authors originally classified the dinosaur as a theropod, a member of a large group of bipedal, mostly carnivorous dinosaurs that includes Tyrannosaurus, but several unpublished opinions beginning in 1995 suggested that the remains actually belonged to a sauropod (probably a titanosaur), a member of a very different group of quadrupedal, herbivorous dinosaurs with long necks and tails. In 2006, the first published reference to Bruhathkayosaurus as a sauropod appeared in a survey of Malagasyvertebrates by David Krause and colleagues.
Until the remains are properly described, the validity of the genus and any size estimates will be questionable. It is possible that the only known remains of Bruhathkayosaurus have been lost to monsoon flooding. Thus the only remaining evidence is likely the very simple and indistinct line-drawings of the bones.
Bruhathkayosaurus was found near the southern tip of India, specifically in the Tiruchirappalli district of Tamil Nadu, to the northeast of Kallamedu village. It was recovered from the rocks of the Kallemedu Formation, which are dated to the Maastrichtian faunal stage of the late Cretaceous period. It lived toward the end of Mesozoic Era, about 70 million years ago. The fossilized remains include hip bones (the ilium and ischium), part of a leg bone (femur), a shin bone (tibia), a forearm (radius) and a tail bone (part of a vertebra, specifically a platycoelous caudal centrum). The remains were originally classified as belonging to a arnosaur. The name chosen, Bruhathkayosaurus, is derived from bruhath (South Indian transliteration of Sanskrit bṛhat "huge, heavy") and kāya (body"), plus the Greek sauros (lizard).
The Bruhathkayosaurus genus has only one known species, Bruhathkayosaurus matleyi. The species is represented by the holotype specimen GSI PAL/SR/20, which was described by Yadagiri and Ayyasami in 1989 (not 1987, as some sources indicate). It was originally classified as a carnosaur (like Allosaurus), of an unknown (incertae sedis) family. It was later recognized as a sauropod.
The original publication described little in the way of diagnostic characteristics and was only supported by a few line drawings. This has led to speculation that the bones might actually be petrified wood, akin to the way the original discoverers of Sauroposeidon initially believed their find to be fossilized tree trunks.
According to the published description, the shin bone (tibia) of Bruhathkayosaurus is 2 m (6.6 ft) long. This is 29 percent larger than the tibia of Argentinosaurus, which is only 1.55 m (5.1 ft) long. The fragmentary femur is similarly huge, across the distal end, it measures 75 cm (2.46 ft), 33% larger than the femur of Antarctosaurus giganteus, which measures 56 cm (1.84 ft)
No total body size estimates for Bruhathkayosaurus have been published, but paleontologists and researchers have posted tentative estimates on the Internet. One early estimate by Mickey Mortimer estimated that Bruhathkayosaurus could have reached 40–44 m (131–144 ft) in length and to have weighed 175–220 tons.However, Mortimer later retracted these estimates, reducing his estimated length of Bruhathkayosaurus to 28–34 m (92–112 ft), and declined to provide a new weight estimate, describing the older mass estimates as inaccurate.In a May 2008 article for the weblog Sauropod Vertebra Picture of the Week, paleontologist Matt Wedel used a comparison with Argentinosaurus and calculated the weight of Bruhathkayosaurus at up to 139 tons.
Amphicoelias,the largest land animal vs possible heaviest Bruhathkayosaurus

From left to right :Camarasaurus lentus, Giraffatitan brancai, Diplodocus hallorum, Spinophorosaurus nigerensis, Glacialisaurus hammeri, Dicraeosaurus hansemanni, Apatosaurus louisae, Daxiatitan binglingi, Brachiosaurus altithorax, Lamplughsaura dharmaramensis, Supersaurus vivianae, Argyrosaurus superbus, Diplodocus carnegii, Apatosaurus ajax, Mamenchisaurus sinocanadorum, Atlasaurus Imelakei, Cetiosaurus oxoniensis, Omeisaurus junghsiensis, Sauroposeidon proteles, Barosaurus lentus,Lusotitan atalaiensis, Amphicoelias altus, Ampelosaurus atacis, Ruyangosaurus giganteus, Paralititan stromeri, Turiasaurus riodevensis, Jobaria tiguidensis, Alamosaurus sanjuanensis, Antarctic titanosaur, Vulcanodon karibaensis, Abydosaurus mcintoshi, Notocolossus gonzalezparejasi, Nigersaurus taqueti, Rhoetosaurus brownei, Brontosaurus parvus, Puertasaurus reuili, Patagotitan mayorum, Diamantinasaurus matildae, Saltasaurus loricatus, Futalognkosaurus dukei, Huanghetitan ruyangensis, Barapasaurus tagorei, Antarctosaurus giganteus, Magyarosaurus dacus, Austrosaurus, Malawisaurus, Savannasaurus elliottorum, Argentinosaurus huinculensis, Dreadnoughtus schrani, Isisaurus colberti and Amargasaurus cazaui.
The sauropods are the gigantic long-necked plant-eating dinosaurs, and include such well-known names as Apatosaurus ("Brontosaurus"), Brachiosaurus, Cetiosaurus, and Diplodocus, as well as numerous other species and genera. They were the largest of all land animals, and among the most successful and long-lived of the dinosaurs.
Among the very first dinosaurs to evolve in the Late Triassic Period, about 230 million years ago (Mya), they became the dominant herbivores by halfway through the late Triassic (during the Norian stage). Their perceived decline in the early Cretaceous is most likely a bias in fossil sampling, as most fossils are known from Europe and North America. Sauropods were still the dominant herbivores in the Gondwanan landmasses, however. The spread of flowering plants (angiosperms) and "advanced" ornithischians, another major group of herbivorous dinosaurs (noted for their highly developed chewing mechanisms), are most likely not a major factor in sauropod decline in the northern continents.Like all non-avian dinosaurs, the sauropodomorphs became extinct 66 Mya, during the Cretaceous–Paleogene extinction event.
The earliest and most basal sauropodomorphs known are Chromogisaurus novasi and Panphagia protos, both from the Ischigualasto Formation, dated to 231.4 million years ago (late Ladinian age of the Middle Triassic according to the ICS; alternately called the early Carnian age of the Late Triassic in the system used by the Geological Society of America).Some studies have found Eoraptor lunensis (also from the Ischigualasto Formation), traditionally considered a theropod, to be an early member of the sauropodomorph lineage, which would make it the most basal sauropodomorph known.
The earliest known species, the small Vulcanodon (6 1/2 metres) dates back to the Sinemurian epoch, while the latest include a number of species such as the small Titanosaurus indicus and Hypselosaurus priscus (lengths about 12 metres), and the large Alamosaurus sanjuanensis (20 metres), were still flourishing right up until the terminal Cretaceous extinction. It was at one time thought on the basis of the North American fossil record) that the sauropods became rare after the Late Jurassic, but in most parts of the world they continued as a common element of the megafauna until the very end of the Cretaceous. Throughout their long history they continued to evolve and branch out new species and families. There are at least three peaks of sauropod diversity in the Late Jurassic, late Early Cretaceous and latest Cretaceous, corresponding to high sea levels and increased speciation resulting from geographical isolation.
Sauropods were long believed to be semi-aquatic swamp wallowers, relying on the bouyancy of water to support their massive bodies. But analysis of their skeletons, in comparison with those of large terrestrial and semi-aquatic animals, and of sedimentation where their fossils have been found, show that sauropods were fully terrestrial:
Not only were sauropods as terrestrial as elephants, but fossil trackways indicate that they lived in herds, again like elephants today. It must have been a truely awesome sight to watch a herd of brontosaurs crossing a Mesozoic floodplain; evoking the same sense of awe, and puniness in one"s own being in comparison, as one would feel when observing whales close up.
The Sauropoda include the largest animals ever to walk on land. These gigantic herbivores reaches lengths of 15 to 25 metres or more (the very largest may have reached 60 metres) and weights of 15 to 30 or even 80 or 150 tonnes.
There are currently eight valid families of sauropods: Vulcanodontidae, Cetiosauridae, Brachiosauridae, Camarasauridae, Diplodocidae, Euhelopodidae, Dicraeosauridae and Titanosauridae.
However, this is certainly an underdestimate of the true diversity of these amazing beasts. Ironically, despite their great size (which would one would think would enable more frequent preservation of remains) the fossil record is quite poor, perhaps due to the large skeletons being skattered before sediment settles to cover them. Most sauropod remains are actually scattered and isolated bones and teeth.
The Sauropods" most defining characteristic was their size. Even the smallest of the species, the dwarf sauropods, grew up to 5 to 6 metres in length, and were counted as some of the largest dinosaurs in their ecosystem. As the largest animals to have ever walked on land, the only competitor in the history of the animal kingdom are the rorqual whales, such as the blue whale.
 While many dinosaurs of different genus had individual variations from their related cousins, the sauropods had little variation between the individual species, possibly due to the forced shared evolution caused by size constraints. However, some members of the genus had individual characteristics, such as the diplodocids, who had extrememly long, whip-like tails, possibly as a defense mechanism against predators. Seismosaurus,the longest of the sauropods, would reach lengths of around 39–52 metres, most likely around 45 metres. Other sauropods, such as the brachiosaurids, were extremely tall, and had high shoulders and extremely long necks used for foraging in the high tree canopies. The Sauroposeidon was most likely the tallest of the sauropods, reaching a height of 18 metres. In terms of today"s animal kingdom, the tallest living animal is the giraffe, which reaches only 4.8 to 5.5 metres in height. Due to their massive height and length, the sauropods were also the largest dinosaurs to walk on the earth. One of heaviest of the class was Argentinosaurus, which most likely weighed approximately 75 to 100 metric tonnes. Other sauropods were of comparable size to the Argentinosaurus, but there is some circumstantial evidence that an even more massive titanosaurian, the Bruhathkayosaurus, weighed a possible 175 to 220 tonnes, although a land animal this size would not have been able to support it"s own weight. By comparison, the largest land animal alive today is the Savannah elephant, which weighs only 10 tonnes.
While many dinosaurs of different genus had individual variations from their related cousins, the sauropods had little variation between the individual species, possibly due to the forced shared evolution caused by size constraints. However, some members of the genus had individual characteristics, such as the diplodocids, who had extrememly long, whip-like tails, possibly as a defense mechanism against predators. Seismosaurus,the longest of the sauropods, would reach lengths of around 39–52 metres, most likely around 45 metres. Other sauropods, such as the brachiosaurids, were extremely tall, and had high shoulders and extremely long necks used for foraging in the high tree canopies. The Sauroposeidon was most likely the tallest of the sauropods, reaching a height of 18 metres. In terms of today"s animal kingdom, the tallest living animal is the giraffe, which reaches only 4.8 to 5.5 metres in height. Due to their massive height and length, the sauropods were also the largest dinosaurs to walk on the earth. One of heaviest of the class was Argentinosaurus, which most likely weighed approximately 75 to 100 metric tonnes. Other sauropods were of comparable size to the Argentinosaurus, but there is some circumstantial evidence that an even more massive titanosaurian, the Bruhathkayosaurus, weighed a possible 175 to 220 tonnes, although a land animal this size would not have been able to support it"s own weight. By comparison, the largest land animal alive today is the Savannah elephant, which weighs only 10 tonnes.The sauropods were herbivorous creatures, and usually were quadrupeds due to their massive girth. They shared a common characteristic of small heads, with large, massive bodies and long, counter-balancing tails. At least some of the species were egg-layers, as evidence has been found that the camarasaurs and titanosaurs laid eggs. Due to their gigantic size, sauropods had thick legs to support their weight, with blunt, balanced feet that ended in five toes.
Amphicoelias fragillimus is a genus of herbivious sauropoddinosaur and is the largest animal to have ever live, bigger than the blue whale.

A mega-sauropod, Amphicoelias fragillimus, is a contender for the largest dinosaur in history. It has been estimated at 58 metres (190 ft) in length and 122,400 kilograms (269,800 lb) in weight. Unfortunately, the fossil remains of this dinosaur have been lost. Amphicoelias (meaning "biconcave", from the Greek amphi: "on both sides", and koilos: "hollow, concave") is a genus of herbivorous sauropod dinosaur that is probably synonymous with the genus Diplodocus. It includes what has sometimes been estimated to be the largest dinosaur specimen ever discovered, originally named "A. fragillimus". Based on surviving descriptions of a single fossil bone, scientists had over the years estimated A. fragillimus to have been the longest known vertebrate at 58 metres (190 ft) in length, with potentially a mass of up to 122.4 tonnes (135 short tons). However, because the only fossil remains were lost at some point after being studied and described in the 1870s, evidence survived only in drawings and field notes. More recent analysis of the surviving evidence, and the biological plausibility of such a large land animal, has suggested that the enormous size of this animal were over-estimates due partly to typographical errors in the original 1878 description.
The type species of Amphicoelias, A. altus, was named by paleontologist Edward Drinker Cope in December 1877 (though not published until 1878) for an incomplete skeleton consisting of two vertebrae, a pubis (hip bone), and a femur (upper leg bone).Cope also named a second species, A. fragillimus, in the same paper. However, all subsequent researchers have considered A. fragillimus to be a synonym of A. altus. Even by 1881 however, it was recognized that A. altus could not be distinguished from other genera, as the features described by Cope were misinterpreted and are widespread.In 1921, Osborn and Mook assigned additional bones to A. altus—a scapula (shoulder blade), a coracoid (shoulder bone), an ulna (lower arm bone), and a tooth. Henry Fairfield Osborn and Charles Craig Mook noted the overall close similarity between Amphicoelias and Diplodocus, as well as a few key differences, such as proportionally longer forelimbs in Amphicoelias than in Diplodocus. The dentition of Amphicoelias is homodont. Its teeth are shaped like long slender cylindrical rods, are spaced apart and project forward towards the front of the mouth. The femur of Amphicoelias is unusually long, slender, and round in cross section; while this roundness was once thought to be another distinguishing characteristic of Amphicoelias, it has since been found in some specimens of Diplodocus as well. A. altus was also similar in size to Diplodocus, estimated to be about 25 m (82 ft) long.While most scientists have used these details to distinguish Amphicoelias and Diplodocus as separate genera, at least one has suggested that Amphicoelias is probably the senior synonym of Diplodocus.
Amphicoelias fragillimus was collected by Oramel Lucas, a fossil collector employed by E. D. Cope, shortly after he was hired by Cope in 1877. Lucas discovered a partial vertebra (the neural arch and spine) of the new sauropod species in Garden Park, north of Caсon City, Colorado, close to the quarry that yielded Camarasaurus. The vertebra was in poor condition, but astonishingly large, measuring 1.5 metres (4.9 ft) up to 2.7 metres (8.9 ft) in height.Lucas shipped the specimen to Cope in the spring or early summer of 1878, and Cope published it as the holotype specimen (catalogue number AMNH 5777) of a new species, A. fragillimus, that August. The name derives from the Latin fragillimus ("very fragile"), referring to the delicateness of the bone produced by very thin laminae (vertebral walls). As revealed in Cope"s notebooks, which he recorded based on Lucas" report on excavation site locations in 1879, the specimen came from a hill south of the Camarasaurus quarry now known as "Cope"s Nipple." While Cope originally wrote that the site belonged to the Dakota Formation (mid-Cretaceous in age), the presence of dinosaurs such as Camarasaurus in the same rocks indicates that they probably belong to the Morrison Formation, which places the age of the site at 150 million years ago in the late Jurassic period, specifically the Tithonian age.
The gigantic bones attributed to A. fragillimus have often been ignored in summaries of the largest dinosaurs partly because, according to various subsequent reports, the whereabouts of both the vertebra and the femur are unknown, and all attempts to locate them have failed.Carpenter, in 2006, presented a possible scenario for the disappearance of the A. fragillimus specimens. As Cope noted in his description, the neural arch bone material was very fragile, and techniques to harden and preserve fossil bone had not yet been invented (Cope"s rival, paleontologist O.C. Marsh, was the first to use such chemicals, in the early 1880s). Carpenter observed that the fossil bones known from the A. fragillimus quarry would have been preserved in deeply weathered mudstone, which tends to crumble easily and fragment into small, irregular cubes. Therefore, the bone may have crumbled badly and been discarded by someone at the American Museum of Natural History (possibly even by Cope himself) soon after he illustrated it in rear view for his paper. Carpenter suggested that this may explain why Cope drew the vertebra in only one view, rather than from multiple angles as he did for his other discoveries.
In 1994, an attempt was made to relocate the original quarry where A. fragillimus and other species had been found, using ground-penetrating radar in an attempt to image bones still buried in the ground. This attempt failed because the fossilized mudstone bones were the same density as the surrounding rock, making it impossible to differentiate the two. A study of the local topography also showed that the fossil-bearing rock strata was severely eroded, and probably was so at the time Lucas made his discovery of A. fragillimus, indicating that a majority of the skeleton was gone by the time the vertebra and femur were recovered.
Cope"s description of A. fragillimus has been met with skepticism, with some researchers noting that there were typographical errors in his measurements. For example, the measurement units are given in (obviously incorrect) centimeters rather than millimeters. Carpenter argued that there is every reason to take Cope at his word, noting that the paleontologist"s reputation was at stake. The discovery took place during the Bone Wars, and Cope"s rival Marsh, who was "ever ready to humiliate" Cope, never called the claims into question. Marsh was known to have employed spies to monitor Cope"s discoveries, and may have even had confirmation.
The enormous size of the Amphicoelias fragillimus bones. Paleontologists Henry Fairfield Osborn and C.C. Mook in 1921, as well as John S. McIntosh in 1998, also accepted Cope"s data without question in published reviews. In 2015, Woodruff and Foster published an analysis of the evidence and circumstances surrounding the publication and interpretation of A. fragillimus.
Producing an estimate of the complete size of A. fragillimus requires scaling the bones of better-known species of diplodocid (a family of extremely long and slender sauropods) in the assumption that their relative proportions were similar. In his original paper, Cope did this by speculating on the size of a hypothetical A. fragillimus femur (upper leg bone). Cope noticed that in other sauropod dinosaurs, specifically A. altus and Camarasaurus supremus, the femora were always twice as tall as the tallest dorsal vertebra, and estimated the size of an A. fragillimus femur to be 3.7 m (12 ft) tall.
In 1994, using the related Diplodocus as a reference, Gregory S. Paul estimated a femur length of 3.1 to 4 metres (10 to 13 ft) for A. fragillimus.The 2006 re-evaluation of A. fragillimus by Ken Carpenter also used Diplodocus as a scale guide, finding a femur height of 4.3 to 4.6 metres (14 to 15 ft).Carpenter went on to estimate the complete size of A. fragillimus, though he cautioned that relative proportions in diplodocids could vary from species to species. Assuming the same proportions as the well-known Diplodocus, Carpenter presented an estimated total length of 58 m (190 ft), which he noted fell within the range presented by Paul in 1994 (40 to 60 metres (130 to 200 ft)). Carpenter pointed out that even the lowest length estimates for A. fragillimus were higher than those for other giant sauropods, such as the diplodocid Supersaurus (32.5 metres (107 ft)), the brachiosaurid Sauroposeidon (34 metres (112 ft)), and the titanosaur Argentinosaurus (30 metres (98 ft)). Carpenter presented more speculative, specific proportions for A. fragillimus (again, based on a scaled-up Diplodocus), including a neck length of 16.75 metres (55.0 ft), a body length of 9.25 metres (30.3 ft), and a tail length of 32 metres (105 ft). He estimated the total forelimb height at 5.75 metres (18.9 ft) and hind limb height at 7.5 metres (25 ft), and the overall height (at the highest point on the back) at 9.25 metres (30.3 ft). By comparison the blue whale, which is on average the longest living creature, reaches 30 metres (98 ft) in length.
 While A. fragillimus was relatively thin, its enormous size still made it very massive. Weight is much more difficult to determine than length in sauropods, as the more complex equations needed are prone to greater margins of error based on smaller variations in the overall proportions of the animal. Carpenter used Paul"s 1994 estimate of the mass of Diplodocus carnegii (11.5 metric tons (11.3 long tons; 12.7 short tons)) to speculate that A. fragillimus could have weighed up to 111.0 metric tons (109.2 long tons; 122.4 short tons).The heaviest blue whale on record weighed about 190 metric tons (190 long tons; 210 short tons), and the heaviest dinosaur known from reasonably good remains, Argentinosaurus, weighed 80 to 100 metric tons (79 to 98 long tons; 88 to 110 short tons), although if the size estimates can be validated, it could still be lighter than Bruhathkayosaurus, which has been estimated to have weighed 139 metric tons (137 long tons; 153 short tons)., but is also known from highly fragmentary remains.
While A. fragillimus was relatively thin, its enormous size still made it very massive. Weight is much more difficult to determine than length in sauropods, as the more complex equations needed are prone to greater margins of error based on smaller variations in the overall proportions of the animal. Carpenter used Paul"s 1994 estimate of the mass of Diplodocus carnegii (11.5 metric tons (11.3 long tons; 12.7 short tons)) to speculate that A. fragillimus could have weighed up to 111.0 metric tons (109.2 long tons; 122.4 short tons).The heaviest blue whale on record weighed about 190 metric tons (190 long tons; 210 short tons), and the heaviest dinosaur known from reasonably good remains, Argentinosaurus, weighed 80 to 100 metric tons (79 to 98 long tons; 88 to 110 short tons), although if the size estimates can be validated, it could still be lighter than Bruhathkayosaurus, which has been estimated to have weighed 139 metric tons (137 long tons; 153 short tons)., but is also known from highly fragmentary remains.Edward Drinker Cope described his finds in two 1878 issues of the American Naturalist, and assigned them to the new genus Amphicoelias. He placed it in a unique family, Amphicoeliidae, though this is now considered a nomen oblitum (forgotten name). The genus is usually assigned to the family Diplodocidae, though some modern analyses have found it at the base of the larger group Diplodocoidea or as a diplodocid incertae sedis (uncertain placement). The first named species in the genus, Amphicoelias altus (holotype specimen AMHD 5764), was discovered by Cope in 1877. But while it is only represented by a partial skeleton, there are enough diagnostic characteristics to provisionally define the genus. A. altus is known from better remains, but is smaller than A. fragillimus. Cope also named a second species in 1878: Amphicoelias latus.
The third named Amphicoelias species, A. fragillimus, was known only from a single, incomplete 1.5 m tall neural arch (the part of a vertebra with spines and processes), either last or second to last in the series of back vertebrae, D (dorsal) 10 or D9. Based only on an illustration published in 1878, this vertebra would have measured 2.7 metres (8.9 ft) tall in life. However, it has been argued that the scale bar in the published description contained a typographical error, and the fossil vertebra was in fact only 1.38 metres (4.5 ft) tall. In addition to this vertebra, Cope"s field notes contain an entry for an "[i]mmense distal end of femur”, located only a few tens of meters away from the giant vertebra. It is likely that this undescribed leg bone belonged to the same individual animal as the neural spine.
In 2007, John Foster suggested that the differences usually cited to differentiate Amphicoelias altus from the more well known Diplodocus are not significant and may be due to individual variation. Foster argued that Amphicoelias is probably the senior synonym of Diplodocus, and that if further research bears this out, the familiar name Diplodocus would need to be abandoned in favor of Amphicoelias, as was the case with Brontosaurus and its senior synonym Apatosaurus.In 2015, Woodruff and Foster reiterated this conclusion, stating that there is only one species of Amphicoelias and that it could be referred to Diplodocus as Diplodocus altus. They considered the name Amphicoelias to be a nomen oblitum.
In his 2006 re-evaluation, Carpenter examined the paleobiology of giant sauropods, including Amphicoelias, and addresses the question of why this group attained such a huge size. He pointed out that gigantic sizes were reached early in sauropod evolution, with very large sized species present as early as the late Triassic Period, and concluded that whatever evolutionary pressure caused large size was present from the early origins of the group. Carpenter cited several studies of giant mammalian herbivores, such as elephants and rhinoceros, which showed that larger size in plant-eating animals leads to greater efficiency in digesting food. Since larger animals have longer digestive systems, food is kept in digestion for significantly longer periods of time, allowing large animals to survive on lower-quality food sources. This is especially true of animals with a large number of "fermentation chambers" along the intestine which allow microbes to accumulate and ferment plant material, aiding digestion. Throughout their evolutionary history, sauropod dinosaurs were found primarily in semi-arid, seasonally dry environments, with a corresponding seasonal drop in the quality of food during the dry season. The environment of Amphicoelias was essentially a savanna, similar to the arid environments in which modern giant herbivores are found, supporting the idea that poor-quality food in an arid environment promotes the evolution of giant herbivores. Carpenter argued that other benefits of large size, such as relative immunity from predators, lower energy expenditure, and longer life span, are probably secondary advantages.
Bruhathkayosaurus
Bruhathkayosaurus ( meaning "huge bodied lizard") is a dinosaur that some researchers claim to have been the largest dinosaur that ever lived. The accuracy of this claim, however, has been mired in controversy and debate. All the estimates are based on Yadagiri and Ayyasami"s 1989 paper, which announced the find.
The authors originally classified the dinosaur as a theropod, a member of a large group of bipedal, mostly carnivorous dinosaurs that includes Tyrannosaurus, but several unpublished opinions beginning in 1995 suggested that the remains actually belonged to a sauropod (probably a titanosaur), a member of a very different group of quadrupedal, herbivorous dinosaurs with long necks and tails. In 2006, the first published reference to Bruhathkayosaurus as a sauropod appeared in a survey of Malagasyvertebrates by David Krause and colleagues.
Until the remains are properly described, the validity of the genus and any size estimates will be questionable. It is possible that the only known remains of Bruhathkayosaurus have been lost to monsoon flooding. Thus the only remaining evidence is likely the very simple and indistinct line-drawings of the bones.
Bruhathkayosaurus was found near the southern tip of India, specifically in the Tiruchirappalli district of Tamil Nadu, to the northeast of Kallamedu village. It was recovered from the rocks of the Kallemedu Formation, which are dated to the Maastrichtian faunal stage of the late Cretaceous period. It lived toward the end of Mesozoic Era, about 70 million years ago. The fossilized remains include hip bones (the ilium and ischium), part of a leg bone (femur), a shin bone (tibia), a forearm (radius) and a tail bone (part of a vertebra, specifically a platycoelous caudal centrum). The remains were originally classified as belonging to a arnosaur. The name chosen, Bruhathkayosaurus, is derived from bruhath (South Indian transliteration of Sanskrit bṛhat "huge, heavy") and kāya (body"), plus the Greek sauros (lizard).
The Bruhathkayosaurus genus has only one known species, Bruhathkayosaurus matleyi. The species is represented by the holotype specimen GSI PAL/SR/20, which was described by Yadagiri and Ayyasami in 1989 (not 1987, as some sources indicate). It was originally classified as a carnosaur (like Allosaurus), of an unknown (incertae sedis) family. It was later recognized as a sauropod.
The original publication described little in the way of diagnostic characteristics and was only supported by a few line drawings. This has led to speculation that the bones might actually be petrified wood, akin to the way the original discoverers of Sauroposeidon initially believed their find to be fossilized tree trunks.
According to the published description, the shin bone (tibia) of Bruhathkayosaurus is 2 m (6.6 ft) long. This is 29 percent larger than the tibia of Argentinosaurus, which is only 1.55 m (5.1 ft) long. The fragmentary femur is similarly huge, across the distal end, it measures 75 cm (2.46 ft), 33% larger than the femur of Antarctosaurus giganteus, which measures 56 cm (1.84 ft)
No total body size estimates for Bruhathkayosaurus have been published, but paleontologists and researchers have posted tentative estimates on the Internet. One early estimate by Mickey Mortimer estimated that Bruhathkayosaurus could have reached 40–44 m (131–144 ft) in length and to have weighed 175–220 tons.However, Mortimer later retracted these estimates, reducing his estimated length of Bruhathkayosaurus to 28–34 m (92–112 ft), and declined to provide a new weight estimate, describing the older mass estimates as inaccurate.In a May 2008 article for the weblog Sauropod Vertebra Picture of the Week, paleontologist Matt Wedel used a comparison with Argentinosaurus and calculated the weight of Bruhathkayosaurus at up to 139 tons.
Тагове:
Encyclopedia Largest Prehistoric Animals...
Encyclopedia Largest prehistoric animals...
Encyclopedia Largest prehistoric animals...
Encyclopedia Largest prehistoric animals...
Encyclopedia Largest prehistoric animals...
Няма коментари
